Suañu
| Suañu | |
|---|---|
|
procesu mental y incidente (es) | |
 | |




Neños pequeños (1-2 años): ente 11 y 14 hores.
Neños n'edá preescolar (3-5 años): ente 10 y 13 hores.
Neños n'edá escolar (6-13 años): ente 9 y 11 hores.
Adolescentes (14-17 años): unes 10,08 hores.
Adultos más mozos (18 a 25 años): ente 7 y 9 hores.
Adultos (26-64 años): ente 7 y 9 hores.
Adultos mayores (de 65 años): ente 7 y 8 hores.
El vocablu suañu (del llatín somnus, que se caltién nos cultismos somníferu, dormiyosu y sonámbulu) designa tantu l'actu de dormir como'l deséu de faelo (tener suañu). Metafóricamente, afírmase qu'una parte del cuerpu durmióse-y a una persona, cuando se pierde o amenorga pasaxeramente la sensibilidá na mesma (parestesia).
Etapes del suañu

Nel adultu, el suañu nocherniegu d'unos ocho hores entamar en 4-5 ciclos d'unos 90-120 minutos, mientres los cualos pásase de la vixilia a la somnolencia (estadiu o fase I), d'ellí al suañu llixeru (fase II), d'ellí al suañu lentu o fondu (fase III) y finalmente al suañu MOR (fase IV). La so distribución estándar nun adultu sanu ye aproximao la siguiente: Fase I, el 5 %; Fase II, el 50 %; Fase III, el 20 % y Fase MOR, el 25 %.
Los estaos y fases del suañu humanu definir según los patrones carauterísticos que se reparen por aciu l'electroencefalograma (EEG), l'electrooculograma (EOG, una midida de los movimientos oculares) y l'electromiograma de superficie (EMG, movimientu de los músculos esqueléticos). El rexistru d'estos parámetros electrofisiológicos pa definir los estaos de suañu y de vixilia denominar polisomnografía.
Estos perfiles describen dos estaos del suañu:
- Suañu ensin movimientos oculares rabiones (suaño non MOR, o NMOR; NREM n'inglés). Nel que pueden estremase tres fases:
- La Fase I del NMOR (fase del suañu llixeru) ye la transición dende la vixilia al suañu; reconocer pola desapaición del patrón regular α (alfa, d'amplitú d'onda alta y regular, de 8 a 13 Hz) y la instauración de la onda θ (theta, d'un patrón d'amplitú baxa y de frecuencia mista, predominantemente nel intervalu de 2 a 7 Hz), con movimientos oculares lentos "en balancín".
- La Fase II del NMOR definir pola apaición de complexos K y de fusos de suañu superpuestos a una actividá de base similar a la del estáu 1. Los complexos K son descargues lentes, negatives (escontra riba) y d'amplitú elevada que se siguen darréu dempués por una deflexión positiva (escontra baxo). Los fusos de suañu son descargues d'alta frecuencia de curtia duración que presenten una amplitú carauterística con xubíes y baxaes. Nun hai actividá ocular y l'EMG da un resultáu similar al estáu 1.
- La Fase III del NMOR (primeramente estremada en fases III y IV) ye la fase de mayor fondura nel suañu, y caracterízase por ser un suañu d'ente 20% a % d'actividá δ (delta, d'amplitú elevada, > 75 microV, y de 0,5 a 2 Hz). Los fusos del suañu pueden persistir y sigue ensin haber actividá ocular. L'actividá del EMG permanez nun nivel amenorgáu y d'altu voltaxe, qu'entiende siquier el 50% del rexistru. La fase III del NMOR denominar suañu d'ondes lentes (SOL), suañu delta o suañu fondu.
- Suañu de movimientos oculares rabiones (MOR; REM n'inglés), o fase IV del suañu, conocíu tamién como suañu paradóxicu, suañu desincronizado o suañu D, carauterizar por un EEG de baxa amplitú y de frecuencia mista similar al de la fase I del NMOR. Prodúcense biltos d'actividá más lenta (3 a 5 Hz) con deflexiones negatives superficiales ("ondes en diente de sierra") que se superponen con frecuencia a esti patrón. El EOG da muestres de movimientos oculares rabiones similares a les que se reparen cuando la persona permanez despierta y colos güeyos abiertos. L'actividá del EMG permanez ausente, un reflexu de l'atonía muscular completa de la paralís motora descendente carauterística d'esti estáu. Nesta etapa preséntense les ensoñaciones (fase onírica). Ye habitual que se presenten de 4 a 6 periodos de suañu MOR mientres la nueche.[2]
-
 Fase 1
Fase 1 -
 Fase 2
Fase 2 -
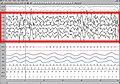 Fase 3
Fase 3 -
 Suaño MOR
Suaño MOR




Necesidaes de suañu y edá
Les necesidaes de suañu varien según la edá. El neñu naciellu duerme casi tol día, con una proporción próxima al 50 % del denomináu suañu «activu», que ye l'equivalente del suañu MOR. A lo llargo de la lactancia los periodos de vixilia son progresivamente más enllargaos y consolídase el suañu de la nueche; amás, la proporción de suañu MOR baxa al 25-30 %, que se va caltener mientres tola vida. De 1-3 años d'edá, el neñu yá solo duerme una o dos siesta. Ente los 4-5 años y l'adolescencia los neños son hipervigilantes, bien pocos duermen pigazu pero tienen un suañu nocherniegu de 9 hores bien estructurado en cinco ciclo o más. Nos individuos nuevos remanez en munchos casos la necesidá fisiolóxica d'un pigazu a metá del día. La necesidá de suañu nun adultu puede bazcuyar ente 5 y 9 hores.
Coles mesmes, varia notablemente l'horariu de suañu ente noctámbulos y madrugadores. En dómines de muncha actividá intelectual o de crecedera o mientres los meses del embaranzu, puede aumentar la necesidá de suañu, ente que l'estrés, l'ansiedá o'l exerciciu físicu practicáu pela tarde pueden menguar la cantidá de suañu. Los estudios efeutuaos n'individuos aisllaos d'influencies esteriores amosaron que l'enclín fisiolóxicu xeneral ye a retrasar llixeramente la fase de suañu con respectu al ciclu convencional de 24 hores y a dormir una curtia pigazu «de mediudía» (Vallina, 2006, 232). Nos vieyos va estazándose el suañu nocherniegu con frecuentes episodios d'espertar y amenórgase enforma'l porcentaxe de suañu en fase IV y non tanto'l de suañu MOR, que se caltién más constante a lo llargo de la vida. Les persones d'edá avanzada tienden a aumentar el tiempu de permanencia na cama. Munches d'elles apigacen fácilmente mientres el día dellos pigazos curtios.
El suañu representa una función vital, por ser imprescindible (el ser humanu nun puede vivir ensin dormir), restauradora (el suañu repara'l cuerpu acaldía), complementaria y fundamental p'asegurar la vixilia (dormir pa poder sentise despiertu a otru día), fisiológicamente necesariu.
Mientres el suañu fondu (Fase III), produzse la restauración física y mientres el suañu MOR la restauración de la función cognitiva (procesu d'aprendizaxe, memoria y concentración).
Función biolóxica del suañu
Eliminación de residuos celulares del celebru
Mientres l'estáu de suañu, el sistema glinfático (equivalente al sistema linfáticu del restu del cuerpu), actívase 10 vegaes más en comparanza al estáu de vixilia, dexando que los residuos de les célules cerebrales esaniciar con mayor eficacia. Mientres el suañu produciría una contraición de les célules cerebrales, creándose asina más espaciu ente elles y collo dexando que'l líquidu cefalorraquídeo circule más fácilmente al traviés del texíu cerebral; llimpiándose asina más llibremente los residuos, tales como la proteína beta-amiloide responsable de la enfermedá d'Alzheimer.[3]
Amás, el cuerpu, mientres el suañu entra n'estáu anabólico, que sirve pa desenvolver numberoses funciones d'arreglu nel nuesu organismu.
Efeutos del suañu
Afitamientu de la memoria
La hipótesis de que'l suañu participa nel afitamientu de la memoria recién foi investigada por aciu cuatro paradigmes:
- Efeutos de la privación del suañu sobre l'afitamientu d'alcordances;
- Efeutos del aprendizaxe sobre'l suañu posentrenamiento;
- Efeutos de la estimulación mientres el suañu sobre'l patrones de suañu y sobre la memoria, #
Reexpresión de los patrones de comportamientu específicu neuronal mientres el suañu posentrenamiento.
Dalgunos d'estos estudios confirmen la idea de que'l suañu ta fondamente implicáu nes funciones de la memoria n'humanos y animales. Sicasí, los datos disponibles entá son demasiáu escasos y n'ocasiones contradictorios pa confirmar o refugar inequívocamente la hipótesis de que l'afitamientu de memories non declarativa y declarativa, respeutivamente, dependan de los procesos de suañu MOR y NMOR. Per otra parte, nun s'atopó nenguna correlación ente la cantidá de suañu que se rexistra nuna especie y la so capacidá intelectual; si'l suañu sirviera pa consolidar la memoria, un gatu que duerme 16 hores diaries tendría de tener una memoria prodixosa, superior a la d'un ser humanu que namái duerme ocho hores. Tamién, persones que nun presenten suañu MOR, por casu, por mancadures traumátiques nel rombencéfalo o debíu al consumu de fármacos, nun tienen nengún problema en consolidar los sos aprendizaxes.[ensin referencies]
Otros estudios más recién comparen el procesu d'ordenamientu de la memoria mientres el suañu col procesu de desfragmentación de la memoria de los ordenadores, dambos escorriendo un mesmu oxetivu de caltenimientu y economía de recursos, preparar pa una meyor disponibilidad operativa de la memoria mientres los momentos de mayor utilidá, como'l tar despiertu o n'actividá.[ensin referencies]
Procesamientu de la memoria
Los científicos demostraron de munches maneres que'l suañu ta rellacionáu cola memoria. Nun estudiu realizáu por Turner, Drummond, Salamat y Brown en persones y animales mientres el suañu demostróse que la memoria de trabayu vese afeutada pola falta de suañu. La memoria de trabayu ye importante porque caltién activa la información pal so posterior procesamientu y sofita les funciones cognitives d'altu nivel, como la toma de decisiones, el razonamientu y la memoria episódica. L'estudiu dexó a 18 muyeres y 22 homes dormir namái 26 minutos pela nueche mientres un periodu de cuatro díes. De primeres los suxetos fueron puestos a prueba en tests cognitivos (folgaos), depués de nuevu dos vegaes al día mientres los cuatro díes de privación de suañu. Na prueba final, la media de memoria de trabayu del grupu priváu de suañu amenorgárase nun 38 per cientu, en comparanza col grupu de control.
Paez que la memoria vese afeutada por distintes etapes del suañu, como'l suañu MOR y el suañu de baxa onda (SBO). Nun estudiu realizáu por Born, Rasch y Gais, tomáronse dellos grupos de suxetos humanos: el grupu de control y el grupu d'ensayu. Encamentóse una xera mental a medianueche (grupu d'ensayu) y otra xera a la fin de la nueche (grupu de control). En cuanto los suxetos dormíen, los hipnogrames marcaben un 23 per cientu de SBO, lo que nos fai saber que mientres la medianueche'l tipu de suañu predominante ye'l SBO. El grupu d'ensayu dio un 16 per cientu más que'l grupu de control (en memoria declarativa), ente que'l grupu de control dio un 25 per cientu más que'l grupu d'ensayu (en memoria de trabayu). Esto indica que la memoria de trabayu (la que más precisamos) ye más alta dempués del periodu de suañu MOR más estensu (escontra'l final de la nueche) y que, sicasí, la memoria declarativa (la que menos precisamos) arriquecer col SBO.
Un estudiu realizáu por Datta, un neurocientíficu de la Escuela de Medicina na Universidá de Boston, sofita indireutamente estes resultancies. Los suxetos fueron 22 aguarones machu. Construyóse una caxa onde un aguarón podía movese llibremente d'un llau a otru. El fondu de la caxa taba fechu con una rejilla d'aceru. Poníase un aguarón na caxa y una activaba una potente lluz acompañada d'un soníu. Dempués de cinco segundos, aplicábase una descarga llétrica nel aguarón. Una vegada aplicada la descarga, l'aguarón podía movese al otru llau de la caxa y evitar la descarga. La duración de la descarga nunca yera de más de 5 segundos. Esto repitióse 30 vegaes na metá de los aguarones. A la otra metá, el grupu de control, fixéron-y lo mesmo pero ensin tener en cuenta la so reacción; dempués de les sesiones, poner a los aguarones n'otra caxa y fixéron-yos rexistros poligráficos llexando seis hores. Esti procesu repitir mientres tres díes. Esti estudiu determinó que, dempués de les sesiones (descargues llétriques), los aguarones pasaben un 25,47 per cientu más tiempu en suañu MOR. Estes pruebes sofiten los resultaos del estudiu de Born y el so equipu, qu'indica una correlación ente'l suañu MOR y la conocencia procedimental.
Tamién se realizó un estudiu alrodiu de la participación de la estimulación de corriente continua na corteza prefrontal p'aumentar la cantidá d'oscilaciones lentes mientres el SWSfe. La estimulación de corriente continua meyora bien de la retención de pareyes de pallabres a otru día, lo que demuestra que'l SWS tien un papel importante nel afitamientu de les alcordances episódiques.[4]
Los diversos estudios suxuren qu'esiste una correlación ente'l suañu y les funciones de la memoria. Los investigadores del suañu de Harvard Saper[5] y Stickgold[6] dixeron qu'una parte esencial de la memoria y del aprendizaxe consiste en que les dendrites de les célules nervioses unvien información a les célules pa faer nueves conexones neuronales. Pa faer esti procesu, les dendrites nun tienen de recibir nengún tipu d'información esterna y por ello suxúrese qu'asocede mientres el suañu.
Árees del encéfalu implicaes nel suañu
- rexón anterior del hipotálamu, área del prosencéfalu basal (suañu);
- rexón posterior del hipotálamu, área del mesencéfalu (vixilia).
Estos dos árees del encéfalu que tán arreyaes na regulación del suañu fueron afayaes, a principios del sieglu XX, primero que surdiera la neurociencia comportamental moderna, pol neurólogu vienés barón Constantin Von Economo. Darréu la implicación d'estos dos árees confirmar por aciu estudios de mancadura n'animales esperimentales (Saper, Chou y Scammell, 2001).
Anque nun s'esclariaron los factores que desencadenen el suañu, delles llinies d'investigación faen pensar que nel encéfalu esisten sustances que lo inducen. Una d'elles podría ser l'adenosina, que s'atropa mientres los periodos d'usu intensu del ATP (adenosín trifosfato) nel sistema nerviosu, y xúnese a receptores específicos, los A1, y inhibe ciertes neurones colinérgicas (lliberadores d'acetilcolina) del sistema d'activación reticular que participen nel espertar. Con éses l'actividá del sistema reticular mientres el suañu ye baxa pol efeutu inhibitorio de la adenosina. La cafeína (del café) y otres metilxantinas, sustances conocíes pol so efeutu de caltener la vixilia, xunir colos receptores A1 y bloquiar, colo que torguen que la adenosina afitar a ellos ya induza el suañu.[7]
Fármacos qu'afecten al suañu
La mayoría de los fármacos qu'inflúin nel suañu pueden clasificase nuna de dos categoríes distintes:
- Hipnóticos: aumenten la cantidá de suañu.
- Antihipnóticos: mengüen la cantidá de suañu.[ensin referencies]
Hai una tercer categoría que cabría introducir, la de los fármacos qu'inflúin sobre la ritmicidad circardiana, siendo'l principal fármacu la melatonina.
Enfermedaes del suañu
- Apnea del suañu. Trestornu en que la persona dexa d'alendar mientres 10s o más, en forma repetitiva, mientres ta dormida, Lo más frecuente ye que se deba a perda del tonu nos músculos de la farinxe, lo cual dexa qu'asoceda'l colapsu de les víes respiratories.[8]
- Narcolepsia. Estáu en que nun se puede tornar el suañu REM mientres los periodos de vixilia. Arriendes d'ello, sobrevienen mientres el día episodios de suañu involuntarios con duración d'unos 15 minutos cada unu.[8]
- Velea familiar fatal. Enfermedá hereditaria bien pocu frecuente.
Aspeutos xenéticos qu'inflúin na duración del suañu
Recién estudios punxeron de manifiestu qu'esisten trés variantes xenétiques rellacionaes cola duración del suañu:
- Variante xenética cercana al xen PAX 8, que produz cambeos con un permediu de 2,6 minutos por apanfilo na duración del suañu. Ye la variante más fuertemente acomuñada a la duración del suañu.
- Variante agües embaxo del xen VRK2, que produz cambeos con un permediu de 2 minutos por apanfilo na duración del suañu.
- Variante agua enriba del xen VRK2, que produz cambeos con un permediu de 1,6 minutos na duración del suañu.[9]
Duración del suañu y enfermedá
Dende va dalgún tiempu, ye conocíu que la duración del suañu ta acomuñada con determinaes enfermedaes metabóliques, tales como obesidá y diabetes tipu 2. Nuevos estudios nesta llinia punxeron de manifiestu qu'esiste una clara correlación xenética ente la curtia duración del suañu y l'índiz de masa corporal (IMC), pero non con diabetes tipu 2; sicasí, esiste una clara correlación xenética ente una duración llarga del suañu, IMC y diabetes tipu 2. Sicasí nun se pudieron establecer fenómenos de causa-efeuto nestos parámetros, por cuenta de los múltiples factores qu'afecten a los mesmos.[9][10][11]
El suañu nos animales

L'actu de suañar foi namái confirmáu nel Homo sapiens. Dellos animales tamién pasen pola fase MOR del suañu, pero la so esperiencia suxetiva ye malo de determinar. Paez que los mamíferos son los animales con mayor probabilidá de suañar por cuenta del so ciclu del suañu similar al humanu. Nel primer llugar de les estadístiques en términos de suañu atopa'l gatu, que pasa un 70 % de la so vida dormiendo y, a midida que avieya, la so etapa de vixilia mengua ostensiblemente.
Los caballos, los coríos y les oveyes pueden dormir de pies o echaos. Sicasí, nun pueden esperimentar suañu MOR mientres tán de pies. L'animal que más tiempu pasa en fase MOR mientres el suañu ye'l armadillu. Les ballenes y los delfines son distintos al restu de los mamíferos: siempres tienen que tar conscientes, yá que precisen salir a la superficie a alendar, solo una parte del so celebru duerme cada vez.[12]
Casi tolos mamíferos que fueron estudiaos amuesen ciclos MOR o NMOR, lo que suxure un mecanismu compartíu al traviés d'especies, con un significáu funcional universal.[13]
-
 Un macacu xaponés dormiendo.
Un macacu xaponés dormiendo. -
 Un dragón de Komodo dormiendo.
Un dragón de Komodo dormiendo. -
 Los felíns son propensos a dormir mientres el día.
Los felíns son propensos a dormir mientres el día.



{kind=link}
Ver tamién
- Suañu
- Fases del suañu
- Inercia del suañu
- Medicina del suañu
- Ritmu circadiano
- Ronquíu
- Trestornu del suañu
Referencies
- ↑ «¿Cuántes hores precisamos dormir según la nuesa edá?». BBC Mundu. Consultáu'l 3 de setiembre de 2016.
- ↑ Sandoval Kingwergs, G. (2002). Factores de riesgu rellacionaos cola velea: Aplicación del Inventariu de Salú, Estilos de Vida y Comportamientu (S.Y.Vi.C.). Tesis de llicenciatura. 219 pp. Méxicu: Facultá de Psicoloxía, UNAM.
- ↑ L. Xie; H. Kang; Q. Xu; M. J. Chen; Y. Liao; T. Meenakshisundaram; J. O’Donnell; D. J. Christensen; J. J. Iliff; T. Takano; R. Deane; M. Nedergaard; C. Nicholson. «Sleep Drives Metabolite Clearance from the Adult Brain.» Science (2013).
- ↑ Marshall et al., 2006, citáu en Walker, M. P. (2009). «The Role of Sleep in Cognition and Emotion». Annals of the New York Academy of Sciences 1156: p. 174.
- ↑ «Hypothalamic regulation of sleep and circadian rhythms». Nature 437 (7063): páxs. 1257-63. 2005. doi:. PMID 16251950.
- ↑ «Sleep-dependent memory consolidation». Nature 437 (7063): páxs. 1272-8. 2005. doi:. PMID 16251952.
- ↑ Tortora, Gerard J. & Reynolds, Sandra. Principios d'anatomía y fisioloxía. Trad. Rubén I. Sánchez M. Oxford University Press. Méxicu. 2002. páxs. 510-511.
- ↑ 8,0 8,1 8,2 Tortora, Gerard J. & Reynolds, Sandra. Principios d'anatomía y fisioloxía. Traducción Rubén I. Sánchez M. Oxford University Press. Méxicu. 2002. páxs. 513.
- ↑ 9,0 9,1 Jones, Samuel Y.; Tyrrell, Jessica; Wood, Andrew R.; Beaumont, Robin N.; Ruth, Katherine S.; Tuke, Marcus A.; Yaghootkar, Hanieh; Hu, Youna et ál. (1 d'agostu de 2016). «Genome-Wide Association Analyses in 128,266 Individuals Identifies New Morningness and Sleep Duration Loci». PLoS genetics 12 (8): páxs. y1006125. doi:. ISSN 1553-7404. PMID 27494321. https://www.ncbi.nlm.nih.gov/pubmed/?term=1.%09Jones,+Samuel+Y.,+et+al.+%22Genome-wide+association+analyses+in+128,266+individuals+identifies+new+morningness+and+sleep+duration+loci.%22+PLoS+Genet+12.8+(2016):+y1006125.. Consultáu'l 19 de febreru de 2017.
- ↑ Cappuccio, Francesco P.; Taggart, Frances M.; Kandala, Ngianga-Bakwin; Currie, Andrew; Peile, Ed; Stranges, Saverio; Miller, Michelle A. (1 de mayu de 2008). «Meta-analysis of short sleep duration and obesity in children and adults». Sleep 31 (5): páxs. 619-626. ISSN 0161-8105. PMID 18517032. https://www.ncbi.nlm.nih.gov/pubmed/?term=2.%09CappuccioFP,TaggartFM,KandalaNB,CurrieA,PeileE,StrangesS,etal.(2008)Meta-analysisof+shortsleepdurationandobesityinchildrenandadults.Sleep+31:619%E2%80%93626.PMID:18517032. Consultáu'l 19 de febreru de 2017.
- ↑ Schmid, Sebastian M.; Hallschmid, Manfred; Schultes, Bernd (1 de xineru de 2015). «The metabolic burden of sleep loss». The Lancet. Diabetes & Endocrinology 3 (1): páxs. 52–62. doi:. ISSN 2213-8595. PMID 24731536. https://www.ncbi.nlm.nih.gov/pubmed/?term=3.%09SchmidSM,HallschmidM,SchultesB(2015)Themetabolicburdenofsleeploss.LancetDiabetes+Endocrinol+3:52%E2%80%9362.doi:10.1016/S2213-8587(14)70012-9PMID:24731536. Consultáu'l 19 de febreru de 2017.
- ↑ Mukhametova LM; Supina AI; Polyakovaa IG (14 d'ochobre de 1977). «Interhemispheric asymmetry of the electroencephalographic sleep patterns in dolphins». Brain Research 134 (3): páxs. 581-584. doi:. PMID 902119.
- ↑ Hobson JA. «Sleep is of the brain, by the brain and for the brain». Nature 437 (7063): páxs. 1254-1256.
Bibliografía
- Bradley, W. G. 2005. páxs. 2021, Neuroloxía Clínica: Diagnósticu y Tratamientu. Madrid: Elsevier España.
- López, J. J. et al. 2002. páxs. 668, DSM-IV-TR: Manual diagnósticu y estadísticu de los trestornos mentales. Barcelona: Masson Eselvier.
- Pocock, G. & Richards, C. D. 2005. páxs. 2005, Fisioloxía humana: la base de la medicina. Barcelona: Elsevier España *
Vallina Ruiloba, J. 2006. páxs. 232, Introducción a la sicopatoloxía y a la psiquiatría. Barcelona: Elsevier España.
Enllaces esternos
 Wikimedia Commons acueye conteníu multimedia sobre Suañu.
Wikimedia Commons acueye conteníu multimedia sobre Suañu. Wikiquote tien frases célebres suyes o que faen referencia a Suañu.
Wikiquote tien frases célebres suyes o que faen referencia a Suañu.