Мітахандрыяльная ДНК
Мітахандрыяльная ДНК (мтДНК) — ДНК, якая знаходзіцца (у адрозненне ад ядзернай ДНК) у мітахондрыях, арганоідах эукарыятычных клетак.
Гены, закадаваныя ў мітахандрыяльнай ДНК, ставяцца да групе плазмагенаў, размешчаных па-за ядром (па-за храмасомай). Сукупнасць гэтых фактараў спадчыннасці, засяроджаных у цытаплазме клеткі, складае плазмон дадзенага віду арганізмаў (у адрозненне ад геному)[1].
Гісторыя адкрыцця
Мітахандрыяльная ДНК была адкрыта Маргіт Нас і Сільвен Нас ў 1963 годзе ў Стакгольмскім універсітэце пры дапамозе электроннай мікраскапіі[2] і, незалежна, навукоўцамі Элен Харлсбрунер, Гансам Тупі і Готфрыдам Шацэм пры біяхімічным аналізе фракцый мітахондрый дрожджаў ў Венскім універсітэце ў 1964 годзе.[3]
Тэорыі ўзнікнення мітахандрыяльнай ДНК
Згодна з эндасімбіятычнай тэорыяй, мітахандрыяльная ДНК паходзіць ад колцавых малекул ДНК бактэрый і таму мае іншае паходжанне, чым ядзерны геном. Цяпер пераважае пункт гледжання, згодна з якой мітахондрыі маюць монафілетычнае паходжанне, гэта значыць былі набыты продкамі эукарыёт толькі аднойчы.
На падставе падабенства ў паслядоўнасцях нуклеатыдаў ДНК бліжэйшымі сваякамі мітахондрый сярод пракарыётаў, якія жывуць цяпер лічаць альфа-пратэабактэрый (у прыватнасці, была прапанавана гіпотэза, што да мітахондрыяў блізкія рыкетсіі). Параўнальны аналіз геномаў мітахондрый паказвае, што ў ходзе эвалюцыі адбывалася паступовае перамяшчэнне генаў продкаў сучасных мітахондрый ў ядро клеткі. Невытлумачальнымі з эвалюцыйнай кропкі гледжання застаюцца некаторыя асаблівасці мітахандрыяльнай ДНК (напрыклад, даволі вялікая колькасць інтронаў, нетрадыцыйнае выкарыстанне трыплетаў і іншыя). З прычыны абмежаванага памеру мітахандрыяльнага геному вялікая частка мітахандрыяльнай бялкоў кадуецца ў ядры. Пры гэтым вялікая частка мітахандрыяльнай тРНК кадуюцца мітахандрыяльным геномам.
Формы і колькасць малекул мітахандрыяльнай ДНК
У большасці вывучаных арганізмаў мітахондрыі ўтрымліваюць толькі кальцавыя малекулы ДНК, у некаторых раслін адначасова прысутнічаюць і кальцавыя і лінейныя малекулы, а ў шэрагу пратыстаў (напрыклад, інфузорый) маюцца толькі лінейныя малекулы[5].
Мітахондрыі млекакормячых звычайна ўтрымліваюць ад двух да дзесяці ідэнтычных копій кальцавых малекул ДНК.[6]
У раслін кожная мітахондрый змяшчае некалькі малекул ДНК рознага памеру, якія здольныя да рэкамбінацыі.
У пратыстаў з атрада кінетапластыдаў (напрыклад, у трыпанасомаў) у адмысловым участку мітахондрыі (кінетапласце) змяшчаецца два тыпы малекул ДНК — ідэнтычныя максі-кольцы (20-50 штук) даўжынёй каля 21 т. п. а. і міні-кольцы (20 000-55 000 штук, каля 300 разнавіднасцяў, сярэдняя даўжыня каля 1000 п. а.). Усе кольцы злучаныя ў адзіную сетку (катенаны), якая руйнуецца і аднаўляецца пры кожным цыкле рэплікацыі. Максі-кольцы гамалагічных мітахандрыяльнай ДНК іншых арганізмаў. Кожнае міні-кальцо змяшчае чатыры падобных кансерватыўных ўчастка і чатыры унікальных гіперварыябельных ўчастка.[7] У міні-кольцах закадаваныя кароткія малекулы накіроўваючых РНК (guideRNA), якія ажыццяўляюць рэдагаванне РНК, транскрыбіруемых з генаў максі-кольцаў.
Устойлівасць мітахандрыяльнай ДНК
Мітахандрыяльная ДНК асабліва адчувальная да актыўных формаў кіслароду, якія генеруюцца дыхальнай ланцугом, у сувязі з іх непасрэднай блізкасцю. Хоць мітахандрыяльная ДНК звязана з бялкамі, іх ахоўная роля менш выказана, чым у выпадку ядзернай ДНК. Мутацыі ў ДНК мітахондрый могуць выклікаць спадчынныя захворванні якіяя перадаюць па мацярынскай лініі. Таксама маюцца дадзеныя, якія паказваюць на магчымы ўклад мутацыі мітахандрыяльнай ДНК у працэс старэння і развіццё узроставых паталогій.[8] У чалавека мітахандрыяльная ДНК звычайна прысутнічае ў колькасці 100-10000 копій на клетку (сперматазоіды і яйцаклеткі з'яўляюцца выключэннем). З множнасцю мітахандрыяльнай геномаў звязаны асаблівасці праявы мітахандрыяльных хвароб — звычайна позні іх пачатак і вельмі зменлівыя сімптомы.
Мітахандрыяльная спадчыннасць
Атрыманне ў спадчыну па мацярынскай лініі
У большасці мнагаклетачных арганізмаў мітахандрыяльная ДНК перадаецца па мацярынскай лініі. Яйкаклетка змяшчае на некалькі парадкаў больш копій мітахандрыяльнай ДНК, чым сперматазоід. У сперматазоідзе звычайна не больш за дзесятак мітахондрый (у чалавека — адна спіральна закручаная мітахондрыя), у невялікіх яйцаклетках марскога яжа — некалькі сотняў тысяч, а ў буйных аацытах жабы — дзесяткі мільёнаў. Акрамя таго, звычайна адбываецца дэградацыя мітахондрый сперматазоіда пасля апладнення[9].
Пры палавым размнажэнні мітахондрыі, як правіла, перадаюцца выключна па мацярынскай лініі, мітахондрыі сперматазоіда звычайна руйнуюцца пасля апладнення. Акрамя таго, большая частка мітахондрый сперматазоіда знаходзяцца ў аснове жгуціка, які пры апладненні часам губляецца. У 1999 годзе было выяўлена, што мітахондрыі сперматазоідаў пазначаныя убіквіцінам (бялком-меткай, якая прыводзіць да разбурэння бацькоўскіх мітахондрый ў зігоце)[10].
Паколькі мітахандрыяльная ДНК не з'яўляецца высокакансерватыўнай і мае высокую хуткасць муціравання, яна з'яўляецца добрым аб'ектам для вывучэння філагеніі (эвалюцыйнага сваяцтва) жывых арганізмаў. Для гэтага вызначаюць паслядоўнасці мітахандрыяльнай ДНК у розных відаў і параўноўваюць іх пры дапамозе спецыяльных камп'ютэрных праграмаў і атрымліваюць эвалюцыйнае дрэва для вывучаных відаў. Даследаванне мітахандрыяльнай ДНК сабакаў дазволіла прасачыць паходжанне сабакаў ад дзікіх ваўкоў[11]. Даследаванне мітахандрыяльнай ДНК у папуляцыях чалавека дазволіла вылічыць «мітахандрыяльную Еву», гіпатэтычную прамаці ўсіх людзей, якія жывуць у наш час.
Атрыманне ў спадчыну па бацькоўскай лініі
Для некаторых відаў паказаная перадача мітахандрыяльнай ДНК па мужчынскай лініі, напрыклад, у мідый[12][13]. Атрыманне ў спадчыну мітахондрый па бацькоўскай лініі таксама апісана для некаторых насякомых, напрыклад, для дразафілы,[14] медоносных пчолаў[15] і цыкадаў.[16]
Існуюць таксама дадзеныя аб мітахандрыялнай перадачы па мужчынскай лініі ў млекакормячых. Апісаны выпадкі такой перадачы ў спадчыну для мышэй,[17][18] пры гэтым мітахондрыі, атрыманыя ад самца, пасля адрываюцца. Такая з'ява паказана для авечак [19] і кланаванай буйной рагатай жывёлы.[20] Таксама апісаны адзіны выпадак, звязаны з бясплоддзем у мужчыны.[21].
Геном мітахондрый
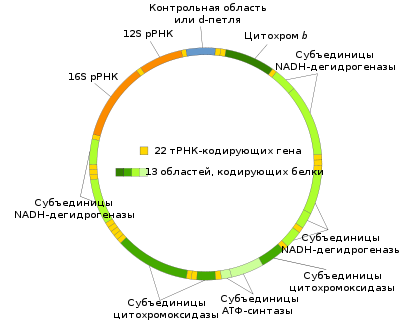
У млекакормячых кожная малекула мтДНК змяшчае 15000-17000 пар аснаванняў (у чалавека 16565 пар нуклеатыдаў — даследаванне скончана ў 1981 годзе[22], паводле іншай крыніцы 16569 пар[23]) і змяшчае 37 генаў — 13 кадуюць бялкі, 22 — гены тРНК, 2 — рРНК (па адным гену для 12S і 16S рРНК). Іншыя шматклеткавыя жывёлы маюць падобны набор мітахандрыяльных генаў, хоць некаторыя гены могуць часам адсутнічаць. Генны склад мтДНК розных відаў раслін, грыбоў і асабліва пратыстаў [24] адрозніваецца больш значна. Так, у жгутыканосца-якабіды Reclinomonas americana знойдзены найбольш поўны з вядомых мітахандрыяльных геномаў: ён змяшчае 97 генаў, у тым ліку 62 гена, кадавальныя бялкі (27 рыбасамальных бялкоў, 23 бялкі, якія ўдзельнічаюць у працы электрон-транспартнага ланцуга і ў акісляльнам фасфарыліраванні, а таксама субадзінак РНК-палімеразы).
Адзін з найбольш маленькіх мітахандрыяльнай геномаў мае малярыйны плазмодый (каля 6.000 п.а., змяшчае два гены рРНК і тры гены, якія кадуюць бялкі).
Нядаўна адкрытыя рудыментарныя мітахондрыі (мітасомы) некаторых пратыстаў (дызентэрыйнай амёбы, мікраспарыдый і лямблій) не ўтрымліваюць ДНК.[25]
Мітахандрыяльныя геномы розных відаў грыбоў утрымліваюць ад 19 431 (дрожджы Schizosaccharomyces pombe) да 100 314 (сардарыёміцэт Podospora anserina) пар нуклеатыдаў[26].
Некаторыя расліны маюць велізарныя малекулы мітахандрыяльнай ДНК (да 25 мільёнаў пар аснаванняў), якія пры гэтым змяшчаюць прыкладна тыя ж гены і ў той жа колькасці, што і меншыя мтДНК. Даўжыня мітахандрыяльнай ДНК можа шырока вар'іравацца нават у раслін аднаго сямейства. У мітахандрыяльнай ДНК раслін маюцца некадуючыя паслядоўнасці якія паўтараюцца.
Геном чалавека змяшчае толькі па адным праматору на кожную камплементарны ланцуг ДНК[22].
Геном мітахондрый чалавека кадуе наступныя бялкі і РНК:
| Бялкі або РНК | Гены |
| NADH-дэгідрагеназа (комплекс I) |
MT-ND1, MT-ND2, MT-ND3, MT-ND4, MT-ND4L, MT-ND5, MT-ND6 |
| Кафермент Q - цытахром c рэдуктаза/Цытахром b (комплекс III) |
MT-CYB |
| цытахром с аксідаза (комплекс IV) |
MT-CO1, MT-CO2, MT-CO3 |
| АТФ-сінтаза | MT-ATP6, MT-ATP8 |
| рРНК | MT-RNR1 (12S), MT-RNR2 (16S) |
| тРНК | MT-TA, MT-TC, MT-TD, MT-TE, MT-TF, MT-TG, MT-TH, MT-TI, MT-TK, MT-TL1, MT-TL2, MT-TM, MT-TN, MT-TP, MT-TQ, MT-TR, MT-TS1, MT-TS2, MT-TT, MT-TV, MT-TW, MT-TY, MT1X |
Асаблівасці мітахандрыяльнай ДНК
Кадавальныя паслядоўнасці (каданом) мітахандрыяльнага геному маюць некаторыя адрозненні ад кадавальных паслядоўнасцяў універсальнай ядзернай ДНК.
Так, кодон AUA кадуе у мітахандрыяльным геноме метыянін (замест ізалейцына ў ядзернай ДНК), кадонам AGA і AGG — тэрмінатарные кадоны (у ядзернай ДНК кадуюць аргінін), кадон UGA у мітахандрыяльным геноме кадуе трыптафан[22].
Калі казаць дакладней, то гаворка ідзе не аб мітахандрыяльнай ДНК, а пра мРНК, якая спісваецца (транскрыбуецца) з гэтай ДНК перад пачаткам сінтэзу бялку. Літара U у пазначэнні кадона пазначае урыдзін, які пры транскрыпцыі РНК гена ў замяняе тымін.
Колькасць генаў тРНК (22 гена) менш, чым у ядзерным геноме з яго 32 генамі тРНК[22].
У чалавечым мітахандрыяльным геноме інфармацыя настолькі сканцэнтраваная, што ў паслядоўнасцях кадавальных мРНК, як правіла, часткова выдаленыя нуклеатыды, якія адпавядаюць 3'-канцавым тэрмінатарным кадонам[22].
Прымяненне
Акрамя выкарыстання пры пабудове розных філагенетычных тэорый, вывучэнне мітахандрыяльнага геному — асноўны інструмент пры правядзенні ідэнтыфікацыі. Магчымасць ідэнтыфікацыі звязана з існуючымі ў мітахандрыяльным геноме чалавека групавымі і нават індывідуальнымі адрозненнямі.
Паслядоўнасць ўчастка гена субадзінке I цытахром с-аксідазы, кадаваных ў мітахандрыяльнай ДНК, шырока выкарыстоўваецца ў праектах, звязаных з ДНК-баркадаваннем жывёл - вызначэннем прыналежнасці арганізма да таму ці іншаму таксону на аснове кароткіх маркераў ў яго ДНК[27][28]. Для баркадавання раслін выкарыстоўваецца пераважна камбінацыя двух маркераў ў пласціднай ДНК[29].
Група Шухрата Міталіпава з цэнтра эмбрыянальных клетак і геннай тэрапіі Арэгонскага ўніверсітэта распрацавала метад замены мітахандрыяльнай ДНК для лячэння спадчынных мітахандрыяльных захворванняў. Цяпер у Вялікабрытаніі пачатыя клінічныя выпрабаванні гэтага метаду, які атрымаў неафіцыйную назву «3-parent baby technique» - «дзіця ад трох бацькоў». Вядома таксама аб нараджэнні ў выніку гэтай працэдуры дзіцяці ў Мексіцы[30].
Зноскі
- ↑ Джинкс Д., Нехромосомная наследственность, пер. с англ., М., 1966; Сэджер Р., Гены вне хромосом, в кн.: Молекулы и клетки, пер. с англ., М., 1966.
- ↑ Nass, M.M. & Nass, S. (1963 at the Wenner-Gren Institute for Experimental Biology, Stockholm University, Stockholm, Sweden): Intramitochondrial Fibers with DNA characteristics (PDF). In: J. Cell. Biol. Bd. 19, S. 593—629. PMID 14086138
- ↑ Ellen Haslbrunner, Hans Tuppy and Gottfried Schatz (1964 at the Institut for Biochemistry at the Medical Faculty of the University of Vienna in Vienna, Австрия): «Deoxyribonucleic Acid Associated with Yeast Mitochondria» (PDF) Biochem. Biophys. Res. Commun. 15, 127—132.
- ↑ Iborra FJ, Kimura H, Cook PR (2004). "The functional organization of mitochondrial genomes in human cells". BMC Biol. 2: 9. doi:10.1186/1741-7007-2-9. PMC 425603. PMID 15157274.
{cite journal}: Папярэджанні CS1: непазначаны свабодны DOI (спасылка) Папярэджанні CS1: розныя назвы: authors list (спасылка) - ↑ Дымшиц Г. М. Сюрпризы митохондриального генома. Природа, 2002, N 6
- ↑ Wiesner RJ, Ruegg JC, Morano I (1992). "Counting target molecules by exponential polymerase chain reaction, copy number of mitochondrial DNA in rat tissues". Biochim Biophys Acta. 183: 553–559. PMID 1550563.
{cite journal}: Папярэджанні CS1: розныя назвы: authors list (спасылка) - ↑ doi:10.1016/j.exppara.2006.04.005 Архівавана 31 сакавіка 2011.
- ↑ Alexeyev, Mikhail F.; LeDoux, Susan P.; Wilson, Glenn L. (July 2004). "Mitochondrial DNA and aging" (PDF). Clinical Science. 107 (4): 355–364. doi:10.1042/CS20040148. PMID 15279618.
- ↑ Ченцов Ю. С. Общая цитология. — 3-е изд. — МГУ, 1995. — 384 с. — ISBN 5-211-03055-9.
- ↑ Sutovsky, P.; et al. (Nov. 25, 1999). "Ubiquitin tag for sperm mitochondria". Nature. 402: 371–372. doi:10.1038/46466. PMID 10586873.
{cite journal}: Непасрэднае ўжыванне et al. у:|author=(даведка); Праверце значэнне даты ў:|year=(даведка) Discussed in [1] Архівавана 19 снежня 2007. - ↑ Vilà C, Savolainen P, Maldonado JE, and Amorin IR (13 June 1997). "Multiple and Ancient Origins of the Domestic Dog". Science. 276: 1687–1689. doi:10.1126/science.276.5319.1687. ISSN 0036-8075. PMID 9180076.
{cite journal}: Папярэджанні CS1: розныя назвы: authors list (спасылка) - ↑ Hoeh WR, Blakley KH, Brown WM (1991). "Heteroplasmy suggests limited biparental inheritance of Mytilus mitochondrial DNA". Science. 251: 1488–1490. doi:10.1126/science.1672472. PMID 1672472.
{cite journal}: Папярэджанні CS1: розныя назвы: authors list (спасылка) - ↑ Penman, Danny (23 August 2002). "Mitochondria can be inherited from both parents". NewScientist.com. Архівавана з арыгінала 11 кастрычніка 2008. Праверана 2008-02-05.
{cite news}: Праверце значэнне даты ў:|date=(даведка) - ↑ Kondo R, Matsuura ET, Chigusa SI (1992). "Further observation of paternal transmission of Drosophila mitochondrial DNA by PCR selective amplification method". Genet. Res. 59 (2): 81–4. PMID 1628820.
{cite journal}: Папярэджанні CS1: розныя назвы: authors list (спасылка) - ↑ Meusel MS, Moritz RF (1993). "Transfer of paternal mitochondrial DNA during fertilization of honeybee (Apis mellifera L.) eggs". Curr. Genet. 24 (6): 539–43. doi:10.1007/BF00351719. PMID 8299176.
- ↑ Fontaine, KM, Cooley, JR, Simon, C (2007). "Evidence for paternal leakage in hybrid periodical cicadas (Hemiptera: Magicicada spp.)". PLoS One. 9: e892. doi:10.1371/journal.pone.0000892.
{cite journal}: Папярэджанні CS1: непазначаны свабодны DOI (спасылка) Папярэджанні CS1: розныя назвы: authors list (спасылка) - ↑ Gyllensten U, Wharton D, Josefsson A, Wilson AC (1991). "Paternal inheritance of mitochondrial DNA in mice". Nature. 352 (6332): 255–7. doi:10.1038/352255a0. PMID 1857422.
{cite journal}: Папярэджанні CS1: розныя назвы: authors list (спасылка) - ↑ Shitara H, Hayashi JI, Takahama S, Kaneda H, Yonekawa H (1998). "Maternal inheritance of mouse mtDNA in interspecific hybrids: segregation of the leaked paternal mtDNA followed by the prevention of subsequent paternal leakage". Genetics. 148 (2): 851–7. PMID 9504930.
{cite journal}: Папярэджанні CS1: розныя назвы: authors list (спасылка) - ↑ Zhao X, Li N, Guo W, et al. (2004). "Further evidence for paternal inheritance of mitochondrial DNA in the sheep (Ovis aries)". Heredity. 93 (4): 399–403. doi:10.1038/sj.hdy.6800516. PMID 15266295.
- ↑ Steinborn R, Zakhartchenko V, Jelyazkov J, et al. (1998). "Composition of parental mitochondrial DNA in cloned bovine embryos". FEBS Lett. 426 (3): 352–6. doi:10.1016/S0014-5793(98)00350-0. PMID 9600265.
- ↑ Schwartz M, Vissing J (2002). "Paternal inheritance of mitochondrial DNA". N. Engl. J. Med. 347 (8): 576–80. doi:10.1056/NEJMoa020350. PMID 12192017.
- ↑ а б в г д Айала Ф. Д. Современная генетика. 1987.
- ↑ http://chemistry.umeche.maine.edu/CHY431/MitoDNA.html Архівавана 13 жніўня 2011.
- ↑ MW Gray, BF Lang, R Cedergren, GB Golding, C Lemieux, D Sankoff, M Turmel, N Brossard, E Delage, TG Littlejohn, I Plante, P Rioux, D Saint-Louis, Y Zhu and G Burger (1998). "Genome structure and gene content in protist mitochondrial DNAs". Nucleic Acids Research. 26: 865–878.
{cite journal}: Папярэджанні CS1: розныя назвы: authors list (спасылка)http://nar.oxfordjournals.org/cgi/content/abstract/26/4/865 - ↑ en:Mitosome#cite note-Leon04-7
- ↑ Дьяков Ю. Т., Шнырева А. В., Сергеев А. Ю. Введение в генетику грибов. — М.: изд. центр «Академия», 2005. — С. 52. — ISBN 5-7695-2174-0.
- ↑ Paul D. N. Hebert, Alina Cywinska, Shelley L. Ball, Jeremy R. deWaard Biological identifications through DNA barcodes(англ.) // Proceedings of the Royal Society of London B: Biological Sciences. — 2003-02-07. — В. 1512. — Т. 270. — С. 313–321. — ISSN 0962-8452. — DOI:10.1098/rspb.2002.2218
- ↑ Živa Fišer Pečnikar, Elena V. Buzan 20 years since the introduction of DNA barcoding: from theory to application // Journal of Applied Genetics. — 2014-02-01. — В. 1. — Т. 55. — С. 43–52. — ISSN 2190-3883. — DOI:10.1007/s13353-013-0180-y
- ↑ CBOL Plant Working Group1, Peter M. Hollingsworth, Laura L. Forrest, John L. Spouge, Mehrdad Hajibabaei A DNA barcode for land plants(англ.) // Proceedings of the National Academy of Sciences. — 2009-08-04. — В. 31. — Т. 106. — С. 12794–12797. — ISSN 0027-8424. — DOI:10.1073/pnas.0905845106
- ↑ Алла Астахова. Тонкая работа - 2. Блог о здравоохранении (22 жніўня 2017).
Спасылкі
- Mitomap — база дадзеных па мітахандрыяльнаму геному чалавека
- Human mtDNA Архівавана 17 сакавіка 2018. — база дадзеных поўных сіквенсаў мтДНК чалавека
- Phylotree М.ван Овена і. М.Кайзера
- Мітахандрыяльная ДНК: дапаможнік для пачынаючага малекулярнага генеалога Архівавана 27 сакавіка 2018.
Гл. таксама
- Мітахондрыя
- Мітахандрыяльная Ева
- Гаплагрупы
- Мітахандрыяльныя захворванні
- Цытаплазматычная мужчынская стэрыльнасць
Тыпы нуклеінавых кіслот | |
|---|---|
| Азоцістыя асновы | |
| Нуклеазіды | Адэназін • Гуаназін • Урыдзін • Тымідзін • Цытыдзін |
| Нуклеатыды | монафасфаты (АМФ • ГМФ • УМФ • ЦМФ) • дыфасфаты (АДФ • ГДФ • УДФ • ЦДФ) • трыфасфаты (АТФ • ГТФ • УТФ • ЦТФ) • цыклічныя (цАМФ • цГМФ • цАДФР) |
| РНК | мРНК • тРНК • рРНК • антысэнсавыя • gRNA • мікра • некадзіруючыя • piРНК • shRNA • малыя інтэрферыруючыя • малыя ядзерныя • малыя ядзеркавыя • тмРНК |
| ДНК | кДНК • Геном • msDNA • Мітахандрыяльная • Хацімодзі-ДНК |
| Аналагі | Гліколь-нуклеінавая кіслата • Замкнёная нуклеінавая кіслата • ПНК • ТНК • Марфаліна |
| Тыпы вектараў | Фазміды • Плазміды • Фаг лямбда • Касміды • Фаг P1 • Фасміды • Штучная бактэрыяльная храмасома • Штучная дражджавая храмасома • Штучная чалавечая храмасома |
Слоўнікі і энцыклапедыі | |
|---|---|
| Нарматыўны кантроль |