Taro
| Taro | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Taro (Colocasia esculenta) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Colocasia esculenta | ||||||||||||
| (L.) Schott |
Der Taro (Colocasia esculenta) ist eine Pflanzenart in der Familie der Aronstabgewächse (Araceae). Er ist als Nutzpflanze für den Menschen von Bedeutung. Die stärkehaltigen Rhizome werden gekocht oder geröstet verzehrt.
Andere Bezeichnungen für Taro sind Wasserbrotwurzel, Kolokasie,[1] Taioba, Eddo, Eddoe, Eddro und Dasheen; in Ghana, Nigeria und dem anglophonen Teil Kameruns Cocoyam, im frankophonen Teil Macabo; auf Hausa Mankani, auf Yoruba Koko oder Lambo und auf Igbo Ede. In alten Nachschlagewerken wie etwa in Pierers Universal-Lexikon findet sich auch die Schreibweise Tarro.
In seinem Werk The Naturalist in Nicaragua (1874, Kapitel V) gibt Thomas Belt den in Nicaragua üblichen Namen Quequisque an. Inzwischen hat sich dort der Name Malanga durchgesetzt, was sich aber eigentlich auf die ähnliche Tannia bezieht. Auch im östlichen Ecuador werden diese beiden unterschiedlichen Gattungen im Volksmund mit dem Namen Papa china bezeichnet. In Indien ist Taro als Arbi und in Japan als Sato-imo (里芋) bekannt.[2] Er wird auch als Elefantenohrpflanze bezeichnet.[3] Auf den Philippinen wird er Gabi genannt.
Auch andere Arten werden mit Taro oder Elefantenohrpflanze bezeichnet – das Riesenblättrige Pfeilblatt (Alocasia macrorrhizos), Tannia (Xanthosoma sagittifolium), Cyrtosperma merkusii und andere Arten der Gattung Colocasia.
Beschreibung


Taro ist eine immergrüne, ausdauernde, krautige Pflanze, die Wuchshöhen zwischen 1 und 2 Metern erreicht. Die Pflanzen bilden ellipsoide bis keulenförmige Rhizome aus, die sowohl vertikal wie auch horizontal im Boden wachsen. Die kartoffelähnlichen Rhizome sind fleischig und haben einen durchschnittlichen Durchmesser von 3 bis 5 Zentimetern. Bei Zuchtformen erreichen die Rhizome Durchmesser bis zu 15 Zentimetern. Sie haben eine braune, raue Schale mit vereinzelten, dünnen Wurzelfäden und ringförmigen Narben, die das weißliche, oft bräunlich bis violett gefleckte, bis violette, stärkereiche „Fleisch“ umgibt. Sie können 3,5 bis 4 Kilogramm, in Einzelfällen bis zu 24,5 kg schwer werden.[4]
Neben den Rhizomen bildet Taro lange, sich horizontal ausbreitende Stolonen aus. Die Stolonen bilden verdickte Nodien.
Die Chromosomenzahl beträgt 2n = 26, 28, 30, 36, 38, 42, 44, 46, 48, 52, 58, 84 oder 116. Die Vielzahl abweichender Chromosomenzahlen liegt in der Tatsache begründet, dass Taro-Chromosomen während der Zellteilung sehr instabil sind und sich unvorhergesehen verhalten. Die häufigsten Chromosomenzahlen sind 2n = 28 oder 42.[5]
Blätter

Jede Pflanze bildet 2, 3 oder mehr, einfache, pfeil- bis herzförmige, exzentrisch zum Spreitengrund hin gestielte Schildblätter (d. h. der Blattstiel sitzt an der Spreitenunterseite an) mit abgerundeten Lappen. Der Sinus reicht meistens nicht bis zum Stielansatz hinunter. Es werden aber auch „normale“ Blätter gebildet, welche den Stielansatz am Spreitengrund haben. Die Blätter können hängend, horizontal, sowie aufrecht mit der Spitze nach oben oder nach unten am Stiel stehen. Manchmal ist der Blattgrund auch oberseits eingefaltet bis muschelförmig.[6] Die weichen, lederartigen, glatten, samtigen Blattspreiten sind unbehaart und grün, dunkelgrün, manchmal auch weißlich oder bläulich-violett panaschiert, an der Unterseite blasser. Es gibt auch Variationen mit blau-violetten oder mit leicht bläulichen, auch mit schwarzblauen und orange-bläulichen Blättern. Die Spreitenoberfläche ist wachsartig und superhydrophob (wasserabweisend).[7] Die Spitze ist spitz- bis stumpfwinklig oder zugespitzt, gelegentlich auch bespitzt.
Die Blattränder sind ganzrandig bis buchtig und oft leicht gewellt oder auch teilweise umgebogen. Die Spreiten messen normalerweise zwischen 10 und 45 Zentimeter in der Länge und 10 bis 35 Zentimeter in der Breite, können aber auch bedeutend größer werden bis 80 mal 60 Zentimeter.[8] Sie werden aufgrund ihrer Größe auch als „Elefantenohren“ bezeichnet.[3]
Die Nervatur ist weißlich bis violettlich, auch bräunlich, es gibt aber auch Variationen mit purpurner Nervatur und Rändern. Es sind drei primäre Hauptnerven vorhanden, einer mittig zur Spitze hin und zwei zu den basalen Lappen. Die sekundären lateralen Adern verlaufen vom basalen Punkt schräg, parallel vorwärts und rückwärts, die tertiären sind netzartig ausgebreitet.
Die normalerweise grünen, nach oben verjüngenden, aufrechten bis auswärts gebogenen, glatten Blattstiele sind basal reitend angeordnet und zwischen 20 und 80 Zentimeter lang, in Extremfällen bis 180 Zentimeter. Der untere Teil, ein Drittel bis die Hälfte, ist obseits auslaufend rinnenförmig, die Rinnenränder sind teils eingerollt. Das Innere der Blattstiele ist schwammartig (sukkulent) mit Luftblasen. Es gibt auch Variationen mit teils orangen sowie rötlichen oder dunkelvioletten Stielen.
Blüten
Wie bei den Aronstabgewächsen üblich, besteht der Blütenstand aus einem Blütenstandschaft, einem Hochblatt (Spatha) und dem Kolben (Spadix).
Es wird zumeist ein einzelner Blütenstand auf einem 15 bis 50 Zentimeter hohen Schaft gebildet. Die lanzettliche Spatha wird zwischen 10 und 40 Zentimeter lang und bis 6 Zentimeter breit. Auf den unteren 4 bis 5,5 Zentimetern bildet sie einen bis 2 Zentimeter durchmessenden grünen Tubus, der sich dann nach der Fruchtreife öffnet. Die Spatha ist cremefarben bis goldgelb und kapuzen-, bootförmig (cymbiform), mit einer eingerollten Spitze. Sie umgibt den etwas vorstehenden Spadix meist rückseitig. Sie kann aber unterschiedlich ausgeformt sein, zurückgebogen, -gerollt, nach hinten hängend, verdrillt, sowie zurückgerollt und vorgebogen oder auch flach nach hinten abstehend.[9][10]
Der herausragende Spadix ist mehr als dreimal so lang wie der Tubus und misst zwischen 12 und 16,5 Zentimeter. Der Spadix teilt sich in einen weiblichen und männlichen Abschnitt, diese werden von einem sterilen getrennt.[8]
Der vom Tubus umwickelte, weibliche Abschnitt an der Basis, ist konisch verdickt und misst bis 3,5 Zentimeter. Die weiblichen fertilen Blüten sind grün, mit weißem Pistill. Der oberständige Fruchtknoten ist unilokular (mit einem Fruchtknotenfach) mit 36 bis 67 Samenanlagen und misst zwischen 1 und 3 Millimeter im Durchmesser. Der untere Abschnitt ist auch mit unfruchtbaren weibliche Blüten durchsetzt. Die sterilen Blüten sind cremefarben bis gelblich und messen etwa 0,5 Millimeter im Durchmesser. Der anschließende, verengte, sterile Abschnitt, mit sterilen weiblichen Blüten, ist zylindrisch und bis etwa 3 Zentimeter lang.
Der männliche Abschnitt im oberen Teil des Spadix ist kegelig und mit 4 bis 6,5 Zentimetern Länge der längste Abschnitt. Die männlichen Blüten sind blassorange mit drei bis sechs verwachsenen Staubblättern. Die Antheren sind zu einem Synandrium verwachsen.
Die unisexuellen Blüten sind nacktblütig (achlamydeisch).
Die weiß-gelbliche bis etwa 4 Zentimeter lange Spitze (Appendix) des Spadix trägt keine Blüten und ist konisch verjüngt. Sie ist normalerweise kürzer als der männliche Abschnitt, kann aber auch länger sein oder ganz fehlen.[11]
Früchte
Die Früchte sind grüne oder rötlich-orange, ellipsoide, etwa 3 bis 5 Millimeter durchmessende Beeren. Die ovoiden, längs gerippten, hellgelblich-bräunlichen Samen sind zwischen 1 und 1,5 Millimeter lang und etwa 0,8 Millimeter breit; es können bis zu 50 Stück pro Beere enthalten sein. Die Tausendkornmasse beträgt nur 0,2 Gramm.[12][13]
Standorte und Verbreitung
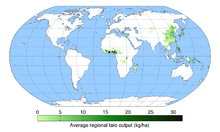
Aufgrund der großen Laubblätter, aus denen viel Feuchtigkeit verdunstet, benötigt Taro sehr viel Wasser. Optimal ist ein Jahresniederschlag zwischen 1500 und 2000 Millimetern. Auch das Substrat sollte sehr feucht sein. Eine Durchschnittstemperatur von 21 °C ist optimal. Die Art ist nicht frostfest. Normalerweise wächst Taro in Höhenlagen bis 1000 Meter; die maximale Höhe, bis zu der Taro noch gedeiht, liegt in Neuguinea bei 2700 Metern. Die Art liebt vollsonnige Standorte und fette Böden mit einem pH-Wert zwischen 5,5 und 6,5 (leicht sauer).
Die größten Tarobestände finden sich in Anpflanzungen auf Feldern oder Wasserfeldern. Wilder Taro bildet oft große Kolonien an feuchten Standorten an Flüssen, Kanälen oder Teichen. Kolonien gibt es aber auch an feuchten Stellen in Wäldern oder in Sumpfgebieten.[14]
Heute ist Taro pantropisch verbreitet, an den meisten Standorten wurde er aber durch den Menschen eingeführt. Forschungen lassen vermuten, dass der ursprüngliche, wilde Taro von der Malaiischen Halbinsel stammt, es aber auch natürliche Vorkommen in Indien gab, wo die Art bereits um 5000 v. Chr. kultiviert wurde. Von dort breitete sich die Art durch den Menschen zunächst in ganz Südostasien, in China und auf Japan aus. Andere Forscher behaupten, dass es auch auf Neuguinea ursprüngliche Taro-Vorkommen gab. In den Pazifik gelangte Taro bereits bei der ersten Besiedlung der Inseln vor bis zu 3500 Jahren.
Kultivierter Taro aus Japan und China gelangte um 100 v. Chr. nach Ägypten, wurde da kultiviert und breitete sich über den Mittelmeerraum aus. Um das Jahr 1 breitete sich die Art entlang der Ostküste Afrikas aus. Von dort gelangte Taro nach Westafrika und vermutlich mit Sklavenschiffen in die Karibik. In Florida gilt Taro als invasives Unkraut.[15]
Nutzung

Taro wird auf der Malaiischen Halbinsel vermutlich seit mehr als 7000 Jahren als Nahrungspflanze kultiviert.[16] Heute ist Taro eine bedeutende Kulturpflanze. Im Jahr 1998 wurden weltweit 6,586 Millionen Tonnen Taroknollen produziert. Die weltweite Anbaufläche lag bei 1,07 Millionen Hektar.[17] Über 80 Prozent der Anbauflächen liegen in Afrika.
Genutzt werden vorwiegend die stärkehaltigen Rhizome der Pflanze. Diese bestehen zu zwei Dritteln aus Wasser und etwa zu einem Drittel aus Kohlenhydraten, zumeist Stärke. Der Proteingehalt beträgt sieben Prozent der Trockenmasse. In den Anbauländern werden aber auch fast alle anderen Pflanzenteile gegessen; besonders Blattstängel und Blätter sind proteinreich (23 Prozent der Trockenmasse). Vor allem die Rhizome enthalten Calciumoxalatkristalle (Raphiden).

In Hawaii ist die dort Kalo genannte Pflanzenart[18] eine der wichtigsten traditionellen Nutzpflanzen. Aus den Rhizomen wird Poi, eine Paste, hergestellt. In einigen Gegenden wird aus den Rhizomen auch Mehl hergestellt; dazu werden die Knollen mit schwefliger Säure behandelt, getrocknet und dann gemahlen.
Auch als Viehfutter ist Taro von Bedeutung; überschüssige oberirdische Pflanzenteile werden in der Regel an Vieh verfüttert, auf Hawaii werden diese Teile auch siliert.
Medizinisch
Die geriebenen Taro-Rhizome (Knollen) werden in der Naturheilkunde in kühlenden Umschlägen zur Behandlung von zum Beispiel Prellungen, Zerrungen, Entzündungen verwendet.[2]
Krankheiten und Schädlinge
Vor allem in Kultur unterliegt Taro gehäuft Angriffen durch Schädlinge. Herauszuheben ist eine Gruppe von Käfern (Coleoptera), die „Taro-Käfer“ genannt werden. Es handelt sich hierbei um Arten aus der Gattung Papuana in der Familie der Blatthornkäfer (Scarabaeidae). Im Speziellen sind dies Papuana woodlarkiana, Papuana biroi, Papuana huebneri und Papuana trinodosa. Die erwachsenen Käfer graben sich zu den Rhizomen vor und fressen diese an. Die angefressenen Rhizome verfaulen in Folge häufig, so dass ein Befall mit Taro-Käfern zu erheblichen Schäden in einer Kultur führen kann. Die weiblichen Käfer legen ihre Eier dicht am Pflanzenstängel ab. Die Larven ernähren sich von Taro-Wurzeln und abgestorbenen Pflanzenteilen, befallen jedoch noch nicht die Rhizome.
Gefürchtet ist auch eine durch Phytophthora colocasiae verursachte Pilzerkrankung, die Taro-Blattfäule genannt wird. Der Pilz verursacht braune Läsionen an den Blättern, aus denen eine gelbliche Flüssigkeit tropft. Binnen 10 bis 20 Tagen stirbt das ganze Blatt ab. Der Befall von Kulturen führt regelmäßig zu Ernteausfällen zwischen 30 und 50 Prozent.
Auch mehrere Pflanzenviren befallen Taro-Kulturen. Bedeutend sind der Taro-Large-Bacilliform-Virus (TLBV), der durch Spitzkopfzikaden (Fulgoromorpha) der Art Tarophagus proserpina übertragen wird, der Taro-Small-Bacilliform-Virus (TSBV), den die Art Planococus citri aus der Familie der Schmierläuse (Pseudococcidae) überträgt, und der Dasheen-Mosaic-Virus (DMV).
Systematik
Colocasia esculenta ist eine von etwa 20 Arten in der Gattung Colocasia. Die Art ist sehr variabel. Sie ist weltweit verbreitet und wurde an ganz verschiedenen Orten durch Zucht oder natürliche Hybridisierung verändert. Dennoch erscheint eine Einteilung der Art in Unterarten und Varietäten nicht sinnvoll.[19]
Durch die starke – auch genetische – Variabilität ist auch eine Einordnung des Taro innerhalb der Gattung Colocasia schwierig. Eine molekulargenetische Untersuchung aus dem Jahr 1998 ergab beispielsweise, dass Colocasia gigantea inzwischen näher mit Arten der Gattungen der Pfeilblätter (Alocasia) und der Goldnarben (Xanthosoma) verwandt ist als mit Colocasia esculenta innerhalb derselben Gattung.[20]
Quellen
Die Informationen im Kapitel Beschreibung entstammen, wenn nicht anders angegeben, den Quellen Li & Boyce 2010 und Thompson 2000. Für die Kapitel Verbreitung, Nutzung, Krankheiten und Schädlinge diente, wenn nicht anders angegeben, Onwueme 1999 als Hauptquelle.
Literatur
- Heng Li, Peter C. Boyce: Colocasia esculenta. In: Flora of China. Band 23. Missouri Botanical Garden Press, St. Louis 2010, ISBN 978-1-930723-99-3, S. 73 f. (englisch, online).
- Sue A. Thompson: Colocasia esculenta. In: Flora of North America. Band 22. Oxford University Press, Oxford 2000, ISBN 978-0-19-513729-3 (englisch, online).
- Inno Onwueme: Taro Cultivation in Asia and the Pacific. Food and Agriculture Organization of the United Nations, Regional Office for Asia and the Pacific, Bangkok 1999 (englisch, PDF; Teil 1, PDF; Teil 2).
- Peter J. Matthews: On the trail of Taro – an exploration of natural and cultural history. In: SENRI Ethnological Studies. Band 88. National Museum of Ethnology, Osaka 2014, ISBN 978-4-906962-17-4 (englisch).
- T. K. Lim: Edible Medicinal and Non Medicinal Plants. Vol. 9, Springer, 2015, ISBN 978-94-017-9510-4, S. 454–492.
- Anton Ivančič, Vincent Lebot: The Genetics and Breeding of Taro. CIRAD, 2000, ISBN 978-2-87614-414-9.
Einzelnachweise
- ↑ Dudenredaktion (Hrsg.): Duden – das große Fremdwörterbuch. 2. Auflage. Bibliographisches Institut, Mannheim 2000, ISBN 3-411-04162-5, S. 719.
- ↑ a b Michio Kushi: Die makrobitische Hausapotheke. 4. Auflage, Ost-West Bund, 1990, ISBN 3-924724-32-6, S. 159–165.
- ↑ a b Ernest Small: Top 100 Food Plants. NRC Press, 2009, ISBN 978-0-660-19858-3.
- ↑ Tipping the scales at 50 pounds, this Kona-grown kalo might set a new world record hawaiinewsnow.com, 3. März 2022, abgerufen am 4. März 2022 (englisch)
- ↑ Onwueme: 1999, S. 4.
- ↑ Anton Ivančič, Vincent Lebot: Descriptors for Taro: (Colocasia Esculenta). International Plant Genetic Resources Institute (IPGRI), 1999, ISBN 92-9043-402-3.
- ↑ Hans J. Ensikat, Petra Ditsche-Kuru u. a.: Superhydrophobicity in perfection: the outstanding properties of the lotus leaf. In: Beilstein J. Nanotechnol. 2, 2011, S. 152–161, doi:10.3762/bjnano.2.19.
- ↑ a b G. J. H. Grubben: Vegetables. Prota, 2004, ISBN 90-5782-147-8, S. 208.
- ↑ W. L. Wagner, D. R. Herbst, S. H. Sohmer: Colocasia. In: Manual of the Flowering Plants of Hawaii. University of Hawai'i Press, Honolulu 1999, ISBN 978-0-8248-2166-1, S. 1356 f.
- ↑ Anton Ivančič, Vincent Lebot: The Genetics and Breeding of Taro. S. 84.
- ↑ S. J. Mayo: Flora of Tropical East Africa: Araceae. Balkema, 1985, ISBN 90-6191-322-5, S. 5.
- ↑ Vincent Lebot: Tropical Root and Tuber Crops. Cabi, 2009, ISBN 978-1-84593-424-8, S. 287.
- ↑ T. K. Lim: S. 461.
- ↑ Thompson: 2000.
- ↑ Steve Lucas: Colocasia esculenta (L.) Schott. In: The Exotic Rainforest. Abgerufen am 11. August 2011 (englisch).
- ↑ Alan Davidson, Helen Saberi, Tom Jaine: The Oxford companion to food. 2. Auflage. Oxford University Press, Oxford 2006, ISBN 978-0-19-280681-9, S. 785 (englisch).
- ↑ Onwueme: 1999, S. 1.
- ↑ taro in Hawaiian Dictionaries, kalo in Hawaiian Dictionaries.
- ↑ Li & Boyce 2010: S. 74.
- ↑ Nguyen Viet Xuan, Yoshino Hiromichi, Tahara Makoto: Phylogenetic Analyses of Taro (Colocasia esculenta (L.) Schott) and Related Species based on Esterase Isozymes. In: Scientific reports of the Faculty of Agriculture, Okayama University. Band 87, Nr. 1, 1998, S. 133–139 (englisch, PDF).
Weblinks
- Colocasia esculenta in der Roten Liste gefährdeter Arten der IUCN 2013.1. Eingestellt von: Nguyen, T.H.T., 2011. Abgerufen am 19. September 2013.