Lípido

En biología y bioquímica, un lípido es una macro biomolécula que es soluble en solventes no polares.[1] Los solventes no polares son típicamente hidrocarburos usados para disolver otras moléculas de lípidos de hidrocarburos que no se disuelven fácilmente (o no se disuelven) en agua, incluyendo ácidos grasos, ceras, esteroles, vitaminas liposolubles (como vitaminas A, D, E y K), monogliceridos, digliceridos, triglicéridos, y fosfolípidos.
Las funciones de los lípidos incluyen almacenamiento de energía, señalización, y actuando como componentes estructurales de membranas celulares.[2][3] Los lípidos tienen aplicaciones en la cosmética y la industria alimenticia así como en la nanotecnología.[4]
Los científicos a veces definen lípidos como hidrófobos y/o moléculas anfilíficas pequeñas; la naturaleza anfilífica de algunos lípidos les permite formar estructuras como vesículas, lisosomas unilamelares/multilamelares, o membranas en un entorno acuoso. Los lípidos biológicos originan enteramente o en parte de dos tipos distintos de subunidades bioquímicas: grupos cetoacil e isopreno.[1] Utilizando esta aproximación, los lípidos se pueden dividir en ocho categorías: ácidos grasos, glicerolípidos, glicerofosfolípidos, esfingolípidos, sacarolípidos, policétidos (derivados de condensación de unidades de cetoacil); lípidos de esterol y lípidos de prenol (derivados de condensación de subunidades de isopreno).[2]
A pesar de que el término "lípido" es a veces utilizado como sinónimo de grasas, las grasas son un subgrupo de los lípidos llamados triglicéridos. Los lípidos también abarcan moléculas como ácidos grasos y sus derivados (incluyendo tri-, di-, monogliceridos, y fosfolípidos), así como otros metabolitos que contienen esterol como colesterol.[5] A pesar de que los humanos y otros mamíferos utilizan varias rutas biosintéticas tanto para romper y sintetizar lípidos, algunos los lípidos esenciales no pueden ser fabricados de este modo y tienen que ser obtenidos de la dieta.
Los lípidos en el cuerpo humano son de crucial importancia para el almacenamiento de energía y el desarrollo de la membrana celular. Los dos tipos principales de lípidos en la sangre son el colesterol y los triglicéridos.
Si los niveles de los lípidos llegan a ser demasiado altos pueden acumularse en las paredes de las arterias hasta formar una placa que puede obstruir el paso de la sangre.
Historia
Los lípidos pueden ser considerados como sustancias orgánicas relativamente insolubles en agua, soluble en solventes orgánicos(alcohol, éter etc.) de hecho o potencialmente relacionados con ácidos grasos y utilizado por las células vivas.
En 1815, Henri Braconnot clasificó a los lípidos (graisses) en dos categorías, suifs (sebo o grasas sólidas) y huiles (aceites fluidos).[6] En 1823, Michel Eugène Chevreul desarrolló una clasificación más detallada, incluyendo aceites, grasas, sebo, ceras, resinas, bálsamos y aceites volátiles (o aceites esenciales).[7][8]
El primer triglicérido sintético fue informado por Théophile-Jules Pelouze en 1844, cuando produjo tributirina al tratar ácido butírico con glicerina en la presencia de ácido sulfúrico concentrado.[6] Varios años más tarde, Marcellin Berthelot, uno de los alumnos de Pelouze, syntetizó triestearina y tripalmitina por reacción del análogo ácido graso con glicerina en la presencia de cloruro de hidrógeno gaseoso a alta temperatura.[5]
En 1827, William Prout reconoció grasa (materias alimentarias "grasosas"), junto con proteína ("albuminosas") y carbohidratos ("sacarina"), como nutrientes importantes para humanos y animales.[8][7]
Por un siglo, los químicos consideraron "las grasas" como lípidos sencillos únicos hicieron de ácidos y glicerol (gliceridos), pero las formas nuevas estuvieron descritas más tarde. Theodore Gobley (1847) descubrió fosfolípidos en huevos de gallina y cerebro mamíferos, él los llamó "lecitinas". Thudichum Descubrió en cerebro humano algunos fosfolípidos (cefalina), glicolípidos (cerebrósido) y esfingolipidos (esfingomielina).[2]
Los términos lipoide, lípido han sido utilizado con significados diversos de autor a autor.[9] En 1912, Rosenbloom y Gies propuso la sustitución de "lipoide" por "lipin".[10] En 1920, Bloor introdujo una clasificación nueva para "lipoides": lipoides simples (grasas y ceras), lipoides compuestos (fosfolipodes y glicolipides), y los derivados de lipoidess (ácidos grasos, alcoholes, esteroles).[11][12]
La palabra lípido, el cual raíces etimológicamente de griegos λίπος, lipos 'grasa', fue introducido en 1923 por el farmacologo francés Gabriel Bertrand.[13] Bertrand incluyó en el concepto no solo las grasas tradicionales (gliceridos), pero también el "lipoides", con una constitución compleja.[2] Incluso aunque la palabra lipide era unánimemente aprobada por la comisión internacional del Société de Chimie Biologique durante la sesión de pleno en el 3 de julio de 1923. La palabra lipide era más tarde anglicismo como lípido debido a su pronunciación ('lɪpɪd). En francés, el sufijo -ide, de Antiguo griego -ίδης (el significado es 'encima de' o 'descendiente de'), es siempre pronunciado (ɪd).
En 1947, T. P. Hilditch Dividió lípidos a "lípidos sencillos", con grasas y ceras (ceras verdaderas, esteroles, alcoholes).
Los lípidos han sido clasificados en ocho categorías por el consorcio Lipid MAPS como sigue:
Ácidos grasos


Los ácidos grasos, o fragmentos de ácidos grasos cuando forman parte de un lípido, son un grupo diverso de moléculas sintetizadas por alargamiento de cadena de acetil-CoA primero con malonil-CoA o grupos metilmalonil-CoA, en un proceso llamado síntesis de ácido graso. Están hechos de un hidrocarburo en cadena que termina con un grupo de ácido carboxílico; este arreglo le confiere a la molécula una terminación polar, hidrófila, y una terminación no polar, hidrófoba que es insoluble en agua. La estructura de los ácidos grasos son de una de las categorías más fundamentales de lípidos biológicos y es generalmente utilizado como bloques de lípidos estructuralmente más complejos. La cadena de carbono, típicamente entre cuatro y veinticuatro carbonos de longitud, pueden ser saturado o insaturados, y pueden estar unidos a grupos funcionales que contienen oxígeno, halógenos, nitrógeno, y azufre. Si un ácido graso contiene un enlace doble, existe la posibilidad de cualquier isomerismo geométrico cis o trans, el cual afecta significativamente la configuración de la molécula. Los enlaces dobles cis causan que la cadena de ácido graso se doble, un efecto que se acentúa cuando hay más dobles enlaces en la cadena. Tres enlaces dobles en el carbono-18 del ácido linolénico, el más abundante ácido graso cadenas de las membranas tilacoides de las plantas, hacen a estas membranas altamente fluidas a pesar de las bajas temperaturas medioambientales, y también hace que el ácido linolénico presente picos agudos en los espectros 13-C NMR de alta resolución de cloroplastos. Esto lo hace jugar un papel importante en la estructura y función de membranas de celulares.[14] La forma más frecuente en que se presentan los ácidos grasos, es la configuración cis, a pesar de que la forma trans existe en algunas grasas y aceites naturales parcialmente hidrogenados.
Ejemplos de biológicamente importantes de ácidos grasos incluyen los eicosanoides, derivados principalmente del ácido ácido araquidónico y ácido eicosapentaenoico, aquello incluye prostaglandinas, leucotrienos, y tromboxanos. Ácido docosahexaenoico el cual también es importante en sistemas biológicos, particularmente en la protección de la vista.[15][16] Otras clases de lípido importantes en la categoría de ácidos grasos son los ésteres grasos y amidas grasas. Los ésteres grasos incluyen importantes intermediarios bioquímicos como los ésteres cerosos, derivados del ácido graso tioester coenzima A, ácido grasos tioester derivados de ACP y carnitinas ácido grasas. Las amidas grasas incluyen N-acil etanolaminas, como el cannabinoide neurotransmisor anandamida.
Glicerolípidos

Los glicerolípidos están compuesto de gliceroles mono-, di-, y trisustituidos, son más conocidos como ácidos grasos triesteres de glicerol, llamados triglicéridos. La palabra "triacilglicérido" es a veces utilizada como sinónimo de "triglicérido". En estos compuestos, los tres grupos hidroxilo de glicerol son esterificados cada uno, típicamente por ácido graso diferente. Como funcionan como un almacenamiento de energía, estos lípidos comprenden la mayor parte de grasa almacenada en los tejidos animales. La hidrólisis de los enlaces éster de los triglicéridos y la liberación de glicerol y los ácidos grasos del tejido adiposo son los pasos iniciales en la metabolización de la grasa.[17]
Subclases adicionales de glcerolípidos están representadas por glicosilgliceroles, los cuales están caracterizados por la presencia de uno o más fragmentos de azúcar unidos a glicerol vía un enlace glucosídico. Ejemplos de las estructuras en esta categoría son el digalactosildiacilgliceroles encontrados en las membranas de las plantas y seminolipidos de células de esperma mamífero.
Glicerofosfolípidos
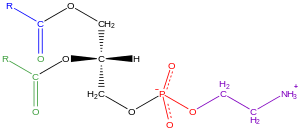
Los glicerofosfolipidos, usualmente son referidos como fosfolípidos (aun así la esfingomielina también se clasifica como fosfolípidos), son ubicuos por naturaleza y son componentes claves de la bicapa lipídica celular, participan en el metabolismo y comunicación celular.[18] El tejido neuronal (incluyendo el cerebro) contiene relativamente cantidades altas de glicerofosfolipidos, y las alteraciones en su composición ha sido asociadas a varios desórdenes neurológicos.[19] Los glicerofosfolipidos se pueden subdividirse en diferentes clases, basados en la naturaleza polar del grupo de la posición sn-3 del glicerol backbone en eucariotas y eubacterias, o la posición sn-1 en el caso de arqueobacterias.
Ejemplos de glicerofosfolipidos los encontramos en las membranas biológicas son fosfatidilcolina (también conocidos como PC, GPCho o lecitina), fosfatidiletaanolamina (PE o GPEtn) y fosfatidilserina (PS o GPSer). Además de servir como componente primario de las membranas celulares y sitios de unión para proteínas intra y intercelulares, algún glicerofosfolipidos en células eucariotas, como fosfatidilinositoles y ácido fosfatídico son precursores o, ellos mismos, derivados de segundos mensajeros de la membrana.[20] Típicamente, uno o ambos de estos los grupos hidroxilo están acilados con ácidos grasos de cadena larga, pero también hay enlaces alquilo y uniones a ácidos grasos de cadena larga, pero hay también alquil-enlazados y 1Z-alquenil-enlazados (plasmalogen) glicerofosfolipidos, así como dialquileter variantes en arqueobacterias.
Esfingolípidos
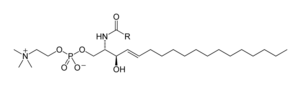
Los esfingolípidos son una familia compleja de compuestos que participación una característica estructural común, una base esfingoide es sintetizada de novo del aminoácido serina y una cadena grasa larga de acyl CoA, convertido entonces a ceramidas, fosfoesfingolipidos, glicoesfingolipidos y otros compuestos.[21] La principal la base esfingoide de los mamíferos es generalmente mencionada como esfingosina. Ceramides (Base N-acyl-esfingoide) son una subclase importante de esfingoides derivados de un enlace amida ácido graso. Los ácidos grasos están típicamente saturados o mono-insaturados con longitudes de cadena de 16 a 26 átomos de carbono.[22]
Los fosfoesfingolipidos importantes de los mamíferos son efingomielinas (ceramida, fosfocolinas), mientras que los insectos contienen principalmente ceramida fosfoetanolaminas y los hongos tienen fosfoinositoles y grupos principales que contienen manosa. Los glicosfingolipidos son una familia diversa de moléculas compuestas de uno o más residuos de azúcar enlazaron vía un enlace glucosídico a la base esfingoide. Ejemplos de estos es el sencillo y complejo glicoesfingolipides como cerebrósidos y gangliósidos.
Esteroles

Los esteroles, como el colesterol y sus derivados, son componentes importantes de lípidos de membrana, junto con los glicerofosfolipidos y esfingomielinas. Otros ejemplos de esteroles son los ácidos biliares y sus conjugados, los cuales en mamíferos son derivados oxidados del colesterol y son sintetizados en el hígado. Los equivalentes en las plantas son los fitosteroles, como el β-sitosterol, estigmasterol, y brassicasterol; este último es también utilizado como biomarcador para el crecimiento del alga. El esterol predominante en las membranas de las células fungi es ergosterol.
Sterols Es esteroides en cuál de los átomos de hidrógeno está sustituido con un hydroxyl grupo, en posición 3 en la cadena de carbono. Han en común con los esteroides igual fusionaron núcleo de cuatro anillos estructura. Los esteroides tienen funciones biológicas diferentes como hormonas y moléculas de señalización. El dieciocho-carbono (C18) los esteroides incluyen el estrogen familia mientras que los #C19 esteroides comprenden el androgens como testosterona y androsterone. La C21 subclase incluye el progestogens así como los glucocorticoides y mineralocorticoids.[23] El secosteroids, comprendiendo varias formas de vitamina D, está caracterizado por cleavage del B anillo de la estructura de núcleo.
Prenoles

Los lípidos de prenol son sintetizados de precursores de unidades de cinco carbonos de isopentenil difosfato y dimetilalil difosfato que se producen principalmente vía ácido mevalónico (MVA). Los isoprenoides sencillos (alcoholes lineales, difosfatos, etc.), se forman por la adición sucesiva de unidades C5, y se clasifican según número de estas unidades de terpeno. Las estructuras grandes que contienen más de 40 carbonos se conocen como politerpenos. Los carotenoides son importantes isoprenoides sencillos que tienen función como antioxidantes y como precursores de vitamina A. Otra clase biológicamente importante de moléculas está ejemplificada por las quinonas e hidroquinonas, los cuales contienen un isoprenoide sujeto a la cola quinonoide núcleo de origen no isoprenoide. La vitamina E y la vitamina K, así como las ubiquinonas, son ejemplos de esta clase. Las procariotas sintetizan poliprenoles (llamados bactoprenoles) en el que la unidad isoprenoide terminal permanece unida al oxígeno, mientras que en los animales poliprenoles (dolicoles) la terminal isoprenoide está reducida.
Sacarolípidos

Los sacarolípidos describen compuestos en los cuales los ácidos grasos están enlazados a un esqueleto de azúcar, la conformación de esta estructura es compatible con bicapas de membrana. En los sacarolípidos, unos sustitutos de monosacárido para el esqueleto glicerol presente en glicerolipidos y glicerofosfolipidos. El sacarolípido más familiar es la glucosamina acilada precursores del Lípido A componente del lipopolisacáridos en bacterias Gram-negativas. La composición típica del lípido A son moléculas disacáridos de glucosamine, los cuales son derivatizados cuando mucho de siete cadenas de graso-acil. El mínimo lipopolisacárido requerido para crecimiento en E. coli es Kdo2-Lípido A, un disacárido hexa-acilado de glucosamina el cual es glicosilado con dos fragmentos de ácido 3-deoxi-D-manno-octulosónico (Kdo).
Policétidos
Los policétidos se sintetizan mediante la polimerización de subunidades de acetilo y propionilo mediante enzimas clásicas, así como enzimas iterativas y multimodulares que comparten características mecánicas con las sintasas de ácidos grasos. Comprenden muchos metabolitos secundarios y productos naturales de origen animal, vegetal, bacteriano, fúngico y marino, y tienen una gran diversidad estructural.[24][25] Muchos policétidos son moléculas cíclicas cuyas cadenas principales a menudo se modifican más mediante glicosilación, metilación, hidroxilación, oxidación u otros procesos. Muchos agentes antimicrobianos, antiparasitarios y anticancerosos de uso común son policétidos o derivados de policétidos, como eritromicinas, tetraciclinas, avermectinas y epotilonas antitumorales.[26]
Funciones biológicas
Membranas
Las células eucariotas presentan la membrana compartimentada-ató orgánulos que lleva a cabo funciones biológicas diferentes. El glycerophospholipids es el componente estructural principal de membranas biológicas, cuando la membrana del plasma celular y las membranas intracelulares de orgánulos; en células animales, la membrana de plasma físicamente separa los componentes intracelulares del extracellular entorno. [cita requerida] El glycerophospholipido es una molécula anfifílica (conteniendo ambas regiones hidrofóbicas e hidrofílicas) aquello contiene un glycerol el núcleo enlazó a dos fatty ácido-derivó "colas" por conexiones de éster y a un "grupo" de cabeza por una conexión de éster del fosfato. [La cita necesitada] Mientras glycerophospholipids es el componente importante de membranas biológicas, otro no-glyceride componentes de lípido como sphingomyelin y sterols (principalmente colesterol en membranas de célula animal) es también encontrado en membranas biológicas.[27] En plantas y algas, el galactosyldiacylglycerols, y sulfoquinovosyldiacylglycerol, los cuales carecen de un grupo de fosfato, son componentes importantes de membranas de chloroplasts y relacionó orgánulos y es los lípidos más abundantes en tejidos fotosintéticos, incluyendo aquellos de plantas más altas, algas y bacterias seguras.[28]
Las membranas tilacoides tienen el componente de lípido más grande de una conformación no-bicapa monogalactosil diglicerido (MGDG), y pocos fosfolípidos; a pesar de esta composición de lípido única, las membranas chloroplasto tilancoides han mostrado contener un lípido bicapa dinámico matricial según lo ha revelado la resonancia magnética y estudios de microscopio electrónico.

Célula
Una membrana biológica es una forma de fase lamelar de una bicapa lipídica. La formación de bicapas lipídicas es un proceso energéticamente preferido cuándo los glicerofosfolipidos descritos arriba se encuentran en un entorno acuoso.[29] Esto se conoce como efecto hidrofóbico. En un sistema acuoso, las cabezas polares de lípidos se alinean hacia el entorno polar, acuoso, mientras las colas hidrofóbicas minimizan su contacto con agua y tienden a agruparse, formando una vesícula; según la concentración del lípido, esta interacción biofísica puede resultar en la formación de micelas, liposomas, o bicapas lipídicas. También se observan otras agregaciones y forman parte del polimorfismo del comportamiento del amfifílo (lípido). El comportamiento de fase es una área de estudio dentro de la biofísica y es el tema de actual [cuando?] búsqueda académica. Micelas y bicapas forma en el medio polar por un proceso conocido como efecto hidrofóbico. Cuando se disuelve una sustancia lipofílica o amfifílica en un entorno polar, las moléculas polares (i.e., agua en soluciones acuosas) se ordenan más alrededor de la sustancia lipofílica disuelta, desde las moléculas polares no pueden formar puentes de hidrógeno a las áreas lipofílicas del amfifílo. Entonces, en un ambiente acuoso, las moléculas de agua forman una jaula de "clatrato" ordenada alrededor de la molécula lipofílica disuelta.
La formación de lípidos en las protocélulas las membranas representa un paso clave en modelos de abiogénesis, el origen de vida.
Almacenamiento de energía
Los triglicéridos, almacenados en tejido adiposo, son una forma importante de almacenamiento de energía tanto en animales como en plantas. Son una fuente importante de energía porque los carbohidratos son estructuras plenamente reducidas. En comparación a glucógeno el cual contribuiría únicamente con la mitad de energía por su masa pura, los carbonos de triglicérido están todos enlazados a hidrógenos, diferentes en carbohidratos.[30] El adipocito, o célula grasa, para la síntesis y descomposición continuas de triglicéridos en animales, con descomposición controlada principalmente por la activación de enzimas sensibles a la hormona lipasa. La oxidación completa de los ácidos grasos proporciona alto contenido calórico, aproximadamente 38 kJ/g (9 kcal/g), comparado con 17 kJ/g (4 kcal/g) por la descomposición de carbohidratos y proteínas. Los pájaros migratorios que tiene que volar distancias largas sin comer usan la energía almacenada de triglicéridos como energía para sus vuelos.[31]
Señalización
Han surgido pruebas que demuestran que la señalización lipídica es una parte vital de la señalización celular.[32][33][34][35] La señalización de los lípidos puede producirse a través de la activación de Proteínas acopladas o receptores nucleares, y se han identificado miembros de varias categorías de lípidos diferentes como moléculas de señalización y mensajeros celulares.[36] Entre ellos se encuentra la esfingosina-1-fosfato, un esfingolípido derivado de la ceramida que es una potente molécula mensajera implicada en la regulación de la movilización del calcio,[37] el crecimiento celular y la apoptosis;[38] el diacilglicerol (DAG) y los fosfatidilinositol fosfatos (PIP), que participan en la activación mediada por el calcio de la proteína quinasa C;[39] las prostaglandinas, que son un tipo de eicosanoide derivado de los ácidos grasos que participa en la inflamación y la inmunidad;[40] las hormonas esteroides como el estrógeno, la testosterona y el cortisol, que modulan una serie de funciones como la reproducción, el metabolismo y la presión arterial; y los oxisteroles como el 25-hidroxicolesterol que son receptores X del hígado agonistas.[41] Se sabe que los lípidos de la fosfatidilserina participan en la señalización para la fagocitosis de las células apoptóticas o de trozos de células. Lo consiguen al quedar expuestos a la cara extracelular de la membrana celular tras la inactivación de flipasas que los colocan exclusivamente en la cara citosólica y la activación de scramblases, que revuelven la orientación de los fosfolípidos. Después de que esto ocurra, otras células reconocen las fosfatidilserinas y fagocitan las células o fragmentos celulares que las exponen.[42]
Otras funciones
Las vitaminas "liposolubles" (A, D, E y K) —que son lípidos a base de isopreno— son nutrientes esenciales que se almacenan en el hígado y en los tejidos grasos, con diversas funciones. Las acilcarnitinas intervienen en el transporte y el metabolismo de los ácidos grasos dentro y fuera de las mitocondrias, donde se someten a la beta oxidación.[43] Los poliprenoles y sus derivados fosforilados también desempeñan importantes funciones de transporte, en este caso el transporte de oligosacáridos a través de las membranas. Los azúcares fosfato de polprenol y los azúcares difosfato de polprenol funcionan en las reacciones de glicosilación extracitoplasmática, en la biosíntesis de polisacáridos extracelulares (por ejemplo, la polimerización del peptidoglicano en las bacterias) y en la N-glicosilación de las proteínas eucariotas.[44][45] Las cardiolipinas son una subclase de glicerofosfolípidos que contienen cuatro cadenas de acilo y tres grupos de glicerol que son particularmente abundantes en la membrana mitocondrial interna.[46][47] Se cree que activan las enzimas implicadas en la fosforilación oxidativa.[48] Los lípidos también forman la base de las hormonas esteroides.[49]
Metabolismo
Los principales lípidos de la dieta de los seres humanos y otros animales son los triglicéridos animales y vegetales, los esteroles y los fosfolípidos de membrana. El proceso del metabolismo de los lípidos sintetiza y degrada las reservas de lípidos y produce los lípidos estructurales y funcionales característicos de los tejidos individuales.
Biosíntesis
En los animales, cuando hay un exceso de oferta de carbohidratos en la dieta, el exceso de carbohidratos se convierte en triglicéridos. Esto implica la síntesis de ácidos grasos a partir de acetil-CoA y la esterificación de los ácidos grasos en la producción de triglicéridos, un proceso llamado lipogénesis.[50] Los ácidos grasos son fabricados por ácidos grasos sintasas que polimerizan y luego reducen las unidades de acetil-CoA. Las cadenas de acilo de los ácidos grasos se extienden mediante un ciclo de reacciones que añaden el grupo acetilo, lo reducen a un alcohol, lo deshidratan a un grupo alqueno y luego lo reducen de nuevo a un grupo alcano. Las enzimas de la biosíntesis de los ácidos grasos se dividen en dos grupos, en los animales y en los hongos todas estas reacciones de la sintasa de los ácidos grasos son llevadas a cabo por una única proteína multifuncional,[51] mientras que en los plástidos de las plantas y en las bacterias son enzimas separadas las que realizan cada paso de la vía.[52][53] Los ácidos grasos pueden ser convertidos posteriormente en triglicéridos que se empaquetan en lipoproteínas y se secretan desde el hígado.
La síntesis de ácidos grasos insaturados implica una reacción de desaturación, por la que se introduce un doble enlace en la cadena de acilo graso. Por ejemplo, en los seres humanos, la desaturación del ácido esteárico por la estearoil-CoA desaturasa-1 produce el ácido oleico. El ácido graso doblemente insaturado ácido linoleico así como el triplemente insaturado ácido alfa-linolénico no pueden ser sintetizados en los tejidos de los mamíferos, por lo que son ácidos grasos esenciales y deben obtenerse de la dieta.[54]
Degradación
La Beta oxidación es el proceso metabólico por el cual los ácidos grasos se descomponen en la mitocondria o en los peroxisomas para generar acetil-CoA. En su mayor parte, los ácidos grasos se oxidan mediante un mecanismo que es similar, pero no idéntico, a una inversión del proceso de síntesis de ácidos grasos. Es decir, se eliminan secuencialmente fragmentos de dos carbonos del extremo carboxilo del ácido después de los pasos de deshidrogenación, hidratación, y oxidación para formar un beta-cetoácido, que se divide por tiolisis. El acetil-CoA se convierte finalmente en ATP, CO2 y H2O mediante el ciclo del ácido cítrico y la cadena de transporte de electrones. Por lo tanto, el ciclo del ácido cítrico puede comenzar en el acetil-CoA cuando la grasa se descompone para obtener energía si hay poca o ninguna glucosa disponible. El rendimiento energético de la oxidación completa del ácido graso palmitato es de 106 ATP.[55] Los ácidos grasos insaturados y de cadena impar requieren pasos enzimáticos adicionales para su degradación.
Nutrición y salud
La mayor parte de la grasa que se encuentra en los alimentos está en forma de triglicéridos, colesterol y fosfolípidos. Algunas grasas de la dieta son necesarias para facilitar la absorción de las vitaminas liposolubles (A, D, E y K) y de los carotenoides.[56] Los seres humanos y otros mamíferos tienen una necesidad dietética de ciertos ácidos grasos esenciales, como el ácido linoleico (un ácido graso omega 6) y el ácido alfa-linolénico (un ácido graso omega-3) porque no pueden sintetizarse a partir de precursores simples en la dieta.[54] Ambos ácidos grasos son ácidos grasos poliinsaturados de 18 carbonos que se diferencian por el número y la posición de los dobles enlaces. La mayoría de los aceites vegetales son ricos en ácido linoleico (cártamo, girasol y maíz). El ácido alfa-linolénico se encuentra en las hojas verdes de las plantas y en algunas semillas, frutos secos y legumbres (en particular lino, colza, nuez y soja).[57] Los aceites de pescado son especialmente ricos en los ácidos grasos omega-3 de cadena larga ácido eicosapentaenoico (EPA) y ácido docosahexaenoico (DHA).[58] Muchos estudios han demostrado los beneficios positivos para la salud asociados al consumo de ácidos grasos omega-3 en el desarrollo infantil, el cáncer, las enfermedades cardiovasculares y diversas enfermedades mentales (como la depresión, el trastorno por déficit de atención e hiperactividad y la demencia).[59][60]
Por el contrario, ahora está bien establecido que el consumo de grasas trans, como las presentes en el aceite vegetal parcialmente hidrogenado, es un factor de riesgo de enfermedad cardiovascular. Las grasas que son buenas para uno pueden convertirse en grasas trans por métodos de cocción inadecuados que dan lugar a una cocción excesiva de los lípidos.[61][62][63]
Unos pocos estudios han sugerido que la ingesta total de grasas en la dieta está relacionada con un mayor riesgo de obesidad[64][65] y diabetes,[66] sin embargo, una serie de estudios muy amplios, incluyendo el Women's Health Initiative Dietary Modification Trial, un estudio de ocho años de duración con 49 000 mujeres, el Nurses' Health Study y el Health Professionals Follow-up Study, no revelaron tales vínculos.[67][68] Ninguno de estos estudios sugirió ninguna conexión entre el porcentaje de calorías procedentes de la grasa y el riesgo de cáncer, enfermedades cardíacas o aumento de peso. The Nutrition Source,[69] un sitio web mantenido por el departamento de nutrición de la T. H. Chan School of Public Health de la Universidad de Harvard, resume las pruebas actuales sobre el efecto de las grasas en la dieta: "Las investigaciones detalladas —muchas de ellas realizadas en Harvard— demuestran que la cantidad total de grasa en la dieta no está realmente relacionada con el peso o la enfermedad".[70]
Véase también
Referencias
- ↑ a b Recherches sur les corps gras d'origine animale. Paris: Levrault. 1823.
- ↑ a b c d Introduction to Lipidomics. Boca Raton: CRC Press. 2012. ISBN 9781466551466.
- ↑ «Sur la nature des corps gras.». Annales de chimie 2 (XCIII): 225-277. 31 de marzo de 1815.
- ↑ «Introduction, History and Evolution.». Lipids. Nutrition and health. Boca Raton: CRC Press. 2015. ISBN 9781482242317.
- ↑ a b C R Séances Acad Sci, Paris, 1853, 36, 27; Ann Chim Phys 1854, 41, 216
- ↑ a b Ann Chim Phys 1844, 10, 434
- ↑ a b «On the ultimate composition of simple alimentary substances, with some preliminary remarks on the analysis of organised bodies in general.». Phil. Trans.: 355-388. 1827.
- ↑ a b «Chronological history of lipid center.». Cyberlipid Center. Archivado desde el original el 13 de octubre de 2017. Consultado el 1 de diciembre de 2017.
- ↑ «Lipids. (Fats, Lipoids. Lipins).». Handbook of Histopathological Techniques (3rd edición). London: Butterworths. 1974. pp. 351-376. ISBN 9781483164793.
- ↑ «Suggestion to teachers of biochemistry. I. A proposed chemical classification of lipins, with a note on the intimate relation between cholesterols and bile salts.». Biochem. Bull. 1: 51-6. 1911.
- ↑ «Outline of a classication of the lipids». Proc. Soc. Exp. Biol. Med. 17 (6): 138-140. 1920. doi:10.3181/00379727-17-75.
- ↑ Lipid Analysis: Isolation, Separation, Identification and Lipidomic Analysis. Bridgwater, England: The Oily Press. 2010. ISBN 9780857097866.
- ↑ «Projet de reforme de la nomenclature de Chimie biologique». Bulletin de la Société de Chimie Biologique 5: 96-109. 1923.
- ↑ Devlin, pp. 193–195.
- ↑ «A Long Lipid, a Long Name: Docosahexaenoic Acid». The Lipid Chronicles. 2 de diciembre de 2011.
- ↑ «DHA for Optimal Brain and Visual Functioning». DHA/EPA Omega-3 Institute.
- ↑ van Holde and Mathews, pp. 630–31.
- ↑ «The Structure of a Membrane». The Lipid Chronicles. 5 de noviembre de 2011. Consultado el 31 de diciembre de 2011.
- ↑ «Glycerophospholipids in brain: their metabolism, incorporation into membranes, functions, and involvement in neurological disorders». Chemistry and Physics of Lipids 106 (1): 1-29. June 2000. PMID 10878232. doi:10.1016/S0009-3084(00)00128-6.
- ↑ van Holde and Mathews, p. 844.
- ↑ «Chapter 14: Sphingolipids: Metabolism and Cell Signaling». Biochemistry of Lipids, Lipoproteins and Membranes (4th edición). Amsterdam: Elsevier. 2002. pp. 373-407. ISBN 978-0-444-51138-6.
- ↑ Devlin, pp. 421–422.
- ↑ Stryer et al., p. 749.
- ↑ Walsh CT (March 2004). "Polyketide and nonribosomal peptide antibiotics: modularity and versatility". Science. 303 (5665): 1805–10. Bibcode:2004Sci...303.1805W. doi:10.1126/science.1094318. PMID 15031493. S2CID 44858908.
- ↑ Caffrey P, Aparicio JF, Malpartida F, Zotchev SB (2008). "Biosynthetic engineering of polyene macrolides towards generation of improved antifungal and antiparasitic agents". Current Topics in Medicinal Chemistry. 8 (8): 639–53. doi:10.2174/156802608784221479. PMID 18473889.
- ↑ Minto RE, Blacklock BJ (July 2008). "Biosynthesis and function of polyacetylenes and allied natural products". Progress in Lipid Research. 47 (4): 233–306. doi:10.1016/j.plipres.2008.02.002. PMC 2515280. PMID 18387369.
- ↑ Stryer et al., pp. 329–331.
- ↑ Heinz E. (1996).
- ↑ Stryer et al., pp. 333–334.
- ↑ «Adipocytes as regulators of energy balance and glucose homeostasis». Nature 444 (7121): 847-53. December 2006. Bibcode:2006Natur.444..847R. PMC 3212857. PMID 17167472. doi:10.1038/nature05483.
- ↑ Stryer et al., p. 619.
- ↑ «Mecanismo modular de inhibición de la señalización Wnt por el factor inhibidor de Wnt 1». Nature Structural & Molecular Biology 18 (8): 886-93. July 2011. PMC 3430870. PMID 21743455. doi:10.1038/nsmb.2081.
- ↑ «Docking of fatty acids into the WIF domain of the human Wnt inhibitory factor-1». Lipids 43 (3): 227-30. March 2008. PMID 18256869. S2CID 31357937. doi:10.1007/s11745-007-3144-3.
- ↑ Wang X (June 2004). "Lipid signaling". Current Opinion in Plant Biology. 7 (3): 329–36. doi:10.1016/j.pbi.2004.03.012. PMID 15134755.
- ↑ Dinasarapu AR, Saunders B, Ozerlat I, Azam K, Subramaniam S (June 2011). "Signaling gateway molecule pages--a data model perspective". Bioinformatics. 27 (12): 1736–8. doi:10.1093/bioinformatics/btr190. PMC 3106186. PMID 21505029.
- ↑ Advertencia de cita: La etiqueta
<ref>con nombreEyster 2007no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreHinkovska-Galcheva 2008no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreSaddoughi 2008no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreKlein 2008no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreBoyce 2008no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreBełtowski 2008no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreBiermann 2013no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Indiveri C, Tonazzi A, Palmieri F (October 1991). «Caracterización del transporte unidireccional de carnitina catalizado por el transportador de carnitina reconstituido de mitocondrias de hígado de rata». Biochimica et Biophysica Acta (BBA) - Biomembranas 1069 (1): 110-6. PMID 1932043. doi:10.1016/0005-2736(91)90110-t.
- ↑ Advertencia de cita: La etiqueta
<ref>con nombreParodi 1979no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreHelenius 2001no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreNowicki 2005no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreGohil 2009no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreHoch 1992no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ «Esteroides». Elmhurst. edu. Archivado desde elmhurst.edu/~chm/vchembook/556steroids.html el original el 23 de octubre de 2011. Consultado el 10 de octubre de 2013.
- ↑ Stryer et al., p. 634.
- ↑ Advertencia de cita: La etiqueta
<ref>con nombreChirala 2004no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreWhite 2005no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreOhlrogge 1997no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ a b Stryer et al., p. 643.
- ↑ Stryer et al., pp. 625-626.
- ↑ Bhagavan, p. 903.
- ↑ Advertencia de cita: La etiqueta
<ref>con nombreRusso 2009no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Bhagavan, p. 388.
- ↑ Advertencia de cita: La etiqueta
<ref>con nombreRiediger 2009no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreGalli 2009no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreMicha 2008no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreDalainas 2008no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreMozaffarian 2007no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreAstrup 2008no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreAstrup 2005no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreAstrup 2008bno puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreBeresford 2005no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ Advertencia de cita: La etiqueta
<ref>con nombreHoward 2006no puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto. - ↑ The Nutrition Source, T. H. Chan School of Public Health, Harvard University
- ↑ Advertencia de cita: La etiqueta
<ref>con nombreurlFatsno puede previsualizarse porque está definida fuera de la sección actual o no está definida en absoluto.
Bibliografía
- Medical Biochemistry. San Diego: Harcourt/Academic Press. 2002. ISBN 978-0-12-095440-7.
- Textbook of Biochemistry: With Clinical Correlations (4th edición). Chichester: John Wiley & Sons. 1997. ISBN 978-0-471-17053-2.
- Biochemistry (6th edición). San Francisco: W.H. Freeman. 2007. ISBN 978-0-7167-8724-2.
- Biochemistry (2nd edición). Menlo Park, California: Benjamin/Cummings Pub. Co. 1996. ISBN 978-0-8053-3931-4.
Enlaces externos
Introducción
- List of lipid-related web sites
- Nature Lipidomics Gateway – Round-up and summaries of recent lipid research
- Lipid Library – General reference on lipid chemistry and biochemistry
- Cyberlipid.org – Resources and history for lipids.
- Molecular Computer Simulations – Modeling of Lipid Membranes
- Lipids, Membranes and Vesicle Trafficking – The Virtual Library of Biochemistry, Molecular Biology and Cell Biology
Nomenclatura
Base de datos
- LIPID MAPS – Comprehensive lipid and lipid-associated gene/protein databases.
- LipidBank – Japanese database of lipids and related properties, spectral data and references.
General
- ApolloLipids – Provides dyslipidemia and cardiovascular disease prevention and treatment information as well as continuing medical education programs
- National Lipid Association – Professional medical education organization for health care professionals who seek to prevent morbidity and mortality stemming from dyslipidemias and other cholesterol-related disorders.
| Control de autoridades |
|
|---|