Axone
Pour les articles homonymes, voir Axone (homonymie).

L'axone, ou fibre nerveuse, est le prolongement du neurone qui conduit le signal électrique du corps cellulaire vers les zones synaptiques. Le long de l'axone, ce signal est constitué de potentiels d'action. Les autres prolongements du neurone sont les dendrites qui conduisent le signal des synapses au corps cellulaire. Les neurones ont le plus souvent un seul axone et plusieurs dendrites. Néanmoins, la terminaison de l'axone est très ramifiée — on parle d'arborisation terminale — ce qui lui permet de contacter plusieurs autres neurones avec la même information.
Au sein du système nerveux central, les axones se regroupent en faisceaux ou tractus, alors que dans le système nerveux périphérique, qui parcourt l'ensemble du corps, ils forment les nerfs.
Chez certaines espèces dont les vertébrés, les axones peuvent être entourés par une gaine de myéline. Celle-ci est synthétisée par les cellules de Schwann dans le système nerveux périphérique et par les oligodendrocytes dans le système nerveux central. La gaine de myéline améliore les propriétés électriques de l'axone et permet une vitesse de conduction plus élevée du signal (jusqu'à 120 m/s).
Les axones permettent également le transport de plusieurs types de protéines, du corps cellulaire où elles sont produites (à partir de l'ADN) vers les synapses où elles assurent diverses fonctions. Des organites telles que les mitochondries sont également transportés dans les axones.
Structure
L'axone prend naissance au niveau du cône d'émergence, zone de jonction avec le corps cellulaire où les potentiels gradués sont sommés et génèrent, si leur somme est (supra-)liminaire, un ou plusieurs potentiels d'action. L'axone se termine par une arborisation terminale. Ses extrémités, ou renflements terminaux, constituent l'élément pré-synaptique de la synapse. Sur une branche terminale, peuvent se rencontrer des « enfilades » de renflements formant des synapses en passant.
L'architecture axonale est maintenue par des éléments du cytosquelette, notamment des faisceaux de microtubules. Ces microtubules sont polarisés avec leur pôle négatif du côté du corps cellulaire et le pôle positif du côté de la partie distale de l'axone. Ainsi, les vésicules ou les organites accrochés à la kinésine se déplacent vers la partie distale de l'axone (transport antérograde) et ceux accrochés à la dynéine se déplacent vers le corps cellulaire (transport rétrograde).
Les plus longs axones du corps humain appartiennent au faisceau pyramidal et issus de la couche V du cortex sensorimoteur et se terminant au sein de la moelle lombosacrée ; les plus longs étant probablement ceux contrôlant les muscles interosseux permettant de déplacer les orteils de gauche à droite[réf. nécessaire].
Les axones font des neurones les plus longues cellules du corps humain.
Les axones ont un diamètre de 1 à 15 μm et la vitesse de conduction des potentiels d'action dépend de ce diamètre (plus le diamètre est gros, plus les potentiels d'action se déplacent vite).
Axone modèle
Les axones les plus épais (à plus fort diamètre) se rencontrent chez les céphalopodes. Notamment chez les calmars géants, des axones de 1 mm de diamètre innervent les muscles du manteau et permettent une contraction rapide et forte de ces muscles pour la nage par réaction. Ces axones ont servi de modèle à deux chercheurs britanniques Hodgkin et Huxley en 1939 pour élucider les mécanismes ioniques du potentiel d'action, les deux chercheurs obtenant le prix Nobel de médecine et de physiologie en 1963 pour cette découverte.
Mise en place
On appelle guidage axonal l'ensemble des processus qui permettent à un axone en croissance d'atteindre spécifiquement sa cellule-cible au cours du développement d'un organisme. À l'extrémité d'un axone en développement se trouve une région aplatie appelée cône de croissance. Cette région émet de fins prolongements appelés filopodes qui sont soutenus par un réseau dynamique de microfilaments d'actine. Le cône de croissance possède des récepteurs spécifiques qui lui confèrent une « sensibilité » pour molécules attractrices ou répulsives (éphrines, nétrine…)[1].
Propagation du potentiel d'action
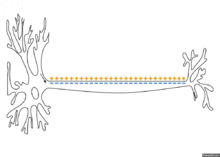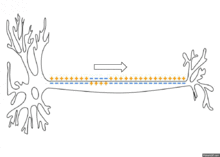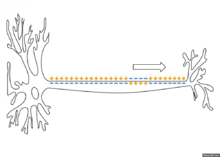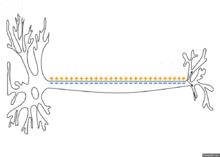



Propagation d'un potentiel décrémentiel
La propagation décrémentielle concerne les dendrites et non l'axone. Il s'agit d'une décroissance exponentielle de l'amplitude des potentiels post-synaptiques activateurs en fonction de la distance parcourue, liée principalement à des courants transmembranaires de fuite.
Propagation de proche en proche (non myélinisé)
Le courant potassique sous-membranaire induit l'ouverture de canaux au sodium voltage-dépendants ce qui entraîne une dépolarisation massive et locale liée à l'entrée d'ions sodium en fonction du gradient électro-chimique. Puis s'activent de manière retardée les canaux potassiques qui laissent sortir les ions potassium en sens inverse repolarisant le neurone. Enfin, une pompe ionique sodium-potassium ATP-asique restaure les concentrations ioniques transmembranaires de repos. Cette séquence se reproduit de proche en proche en raison de la juxtaposition serrée de ces canaux.
Le signal est dans ce cas lent (de l'ordre de 1 à 10 m/s), mais peut parcourir de longues distances (plusieurs mètres dans certains cas chez l'humain).
Propagation saltatoire (myélinisé)
Ce type de propagation est très particulier, mettant en jeu un deuxième type de cellule (oligodendrocyte ou cellule de Schwann). En effet, des gaines de myéline entourent l'axone. C'est un isolant. À ce niveau, rien n'entre, rien ne sort. Ainsi les charges positives se diluent, jusqu'à atteindre une pompe sodique voltage-dépendant entre deux gaines de myéline (ou nœud de Ranvier). Cette pompe va régénérer le potentiel d'action, qui a légèrement diminué à la suite de la dilution. On retrouve encore des pompes et canaux ioniques au niveau du nœud de Ranvier, mais leur nombre est relativement faible.
Le signal est ici très rapide (environ 50 m/s chez l'homme), et arrive à l'extrémité avec la même intensité qu'à l'origine.
Il existe plusieurs types de fibre myélénisées à des niveaux différents et possédant donc une vitesse de transmission du signal différente.
Les fibres B sont des fibres nerveuses moyennement myélénisées, c'est-à-dire moins que les fibre A et plus que les fibres C. Leur vitesse de conduction est entre 3 et 14 m/s. Ce sont généralement des fibres sensitives viscérales et des fibres automatiques pré-ganglionnaires.
Références
- Patrick Pla, « Croissance et guidage axonal », sur Biologie Cellulaire et Génétique du Développement, (consulté le )
Voir aussi
Articles connexes
Liens externes
- Ressources relatives à la santé :
- Notices dans des dictionnaires ou encyclopédies généralistes :