根

根 (ね、root) は、葉や茎とともに、維管束植物 (広義のシダ植物と種子植物) の体を構成する器官の1つである。ふつう地中にあって植物体を基質に固定し、地上部を支えるとともに (図A)、水や無機養分を吸収する役割を担っている (→#根の機能)。


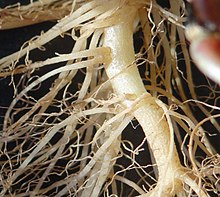
根は先端成長を行い (基本的に先端部だけで細胞分裂を行う)、それを司る根端分裂組織は根冠とよばれる保護構造で覆われている (→#根端)。根は外側から表皮、皮層、中心柱からなり、先端付近の表皮からは根毛とよばれる細長い突起が生じ、吸水面積を広げ、根を土壌に密着させる (図B) (→#内部構造)。中心柱内には吸収した水や無機栄養分を茎や葉に運ぶ木部と葉からの光合成産物が通る師部が放射状に配置しており (放射中心柱)、中心柱は外部との物質連絡を調節する内皮で囲まれている。多くの維管束植物では、内部で形成された新たな根が外側を突き破って伸びることで内生的に側方分枝するが (図C)、小葉植物では外生的に二又分枝する (→#分枝)。胚の時期 (種子の中など) に形成された幼根に由来する根を定根、二次的に茎から生じたものなどそれ以外の根を不定根とよぶ (→#定根と不定根)。木本植物 (木) では、茎と同様に根も維管束形成層による二次成長を行う (→#一次成長と二次成長)。
根はふつう地中にあるが、地上部にあって呼吸や支持、付着、光合成など特殊な機能を担っていることがある (→#さまざまな根)。根はふつう菌根菌と共生して菌根を形成しており、マツタケやトリュフは菌根菌の例である (→#他生物と共生した根)。窒素固定を行う細菌が根に共生している例もある (シロツメクサなど)。また寄生植物は、根を使って他の植物に寄生している。根の中には、食用 (ダイコン、サツマイモ、ニンジンなど) や薬用 (高麗人参やハシリドコロなど) とされるものがある (→#人間との関わり)。
上記のように根は基本的に維管束植物の器官を意味するが、コケ植物や藻類、固着動物など他の生物群がもつ類似の構造を便宜的に根とよぶこともある[2]。以下では維管束植物の器官である根について解説する。
構造
維管束植物の生活環において主要な世代である胞子体 (ゲノムを2セットもち、減数分裂によって胞子を形成する体) は、茎と葉 (シュートとしてまとめられることもある) および根からなる[3][4][5]。例外的に、マツバラン類 (ハナヤスリ亜綱)[6] やコイチヨウラン、オニノヤガラ (ラン科) など菌根菌に大きく依存している植物、サンショウモ属 (薄嚢シダ類)[7] やミジンコウキクサ (サトイモ科) など一部の浮水植物、エアープランツであるサルオガセモドキ (パイナップル科) などは少なくとも成熟した状態では根をもたない[8]。
根端

根はふつう細長い軸状の構造であり、先端成長する[9][10]。根の先端部分は根端 (root apex) とよばれる[10][11]。根端の中には根端分裂組織 (root apical meristem, RAM) とよばれる分裂組織が存在し、活発な細胞分裂を行っている[4][12][13] (図1a)。茎のシュート頂分裂組織とは異なり、根端分裂組織の先端側は根冠 (root cap) とよばれる多細胞層の柔組織によって覆われている[14][4][15] (図1a)。根端分裂組織は先端側に根冠を、基部側に新たな根の組織を作り出して成長していく。
根は土壌中を伸びていくため、先端表面にある根冠の細胞は次第にはがれ落ちていくが (ふつう1個の根冠細胞の寿命は1〜9日ほど)、根端分裂組織によって内側から順次新たな根冠細胞が供給され、根冠には一定量の細胞が維持されている[14][15][16][17] (図1a)。根冠の細胞はムシゲル (粘質ゲル[15]、mucigel) を分泌し、根端を保護すると共に根を伸長しやすくする[15][17]。根はふつう正の重力屈性 (屈地性; 下方へ伸びる性質) を示すが、根冠中央基部付近の細胞 (平衡細胞) 内でアミロプラスト (光合成能を欠き、デンプン粒を多く含む色素体) が沈降することが重力方向の感知に関わっていると考えられている[18]。
根端分裂組織からは基部側へも新たな細胞が付加され、これが拡大伸長し、それに伴い組織分化していくことで根が伸長していく[4][19][20]。根端分裂組織から基部側へつくられた組織は、外側から前表皮 (protoderm)、基本分裂組織 (ground meristem)、前形成層 (procambium) とよばれ、これがそれぞれ表皮、皮層、中心柱へと分化する[4][12][13]。構成する細胞の状態に応じて、根は先端側から大まかに (重なりながら) 分裂帯 (細胞分裂帯[4]、分裂領域 meristematic zone[21])、伸長帯 (伸長領域 elongation zone[21])、成熟帯 (分化帯[4]、分化領域 differentiation zone[21]) に分けられる[22]。根端分裂組織を含む部分が分裂帯であり、細胞数が増加していく。ふつう根端から数 mm のところが伸長帯であり、細胞が拡大伸長している[4]。根の伸長はこの部分で最も活発であり、細胞はときに10倍以上に伸長する。細胞はこの部分で分化し始め、やがて成熟帯において細胞分化が完了する[4]。
内部構造
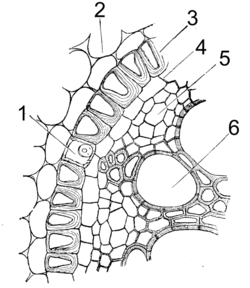



根は、基本的に外側から表皮、皮層、中心柱 (維管束柱[4]) からなる[15][23] (図2a, b)。ただし活発な二次成長を行った根ではほとんどが二次維管束からなり、表面は周皮で覆われている (下記)。
根の表面はふつう1層の細胞からなる表皮 (epidermis; 根の表皮は特に rhizodermis とも表記される[24]) によって囲まれている[15][16][23] (図2a, b)。地上部のシュート (茎や葉) とは異なり、地中の根の表皮ではクチクラ層があまり発達しておらず (そのため吸水できる)、また気孔も存在しない[25]。根の表皮は、根端分裂組織からやや離れたところで根毛 (root hair) を形成する[10][15][26][27]。シロイヌナズナ (アブラナ科) などでは、不等分裂によって形成された小型の根毛形成細胞 (原根毛、trichoblast) が伸長して根毛となる[14][16][24][26]。根毛は直径 10 µm ほどであり、ふつう短命であるが、半年以上残存するものもある (宿存根毛)[14][15][26]。根毛の存在は、土壌粒子との密着や吸水する根の表面積の増大に寄与すると考えられている[10][15][26]。
表皮の内側には、皮層 (cortex) が存在する[15][23][28]。皮層は主に柔細胞からなり、デンプンなどの養分貯蔵に重要な役割を果たすことがある。また根の皮層には大きな細胞間隙が存在することが多く (特に水生植物など)、根の呼吸におけるガス交換に有用であると考えられている[9][12][15][29] (図2a)。皮層の最外層 (表皮のすぐ内側) にある1〜数層は、下皮 (hypodermis) とよばれる[9]。下皮はときにスベリンやリグニンを沈着して細胞壁が厚化し、またカスパリー線が存在することがあり、このような下皮は外皮 (exodermis) とよばれる[9][15][23][30]。外皮は、はがれ落ちた表皮に代わって根の保護構造となる。一方、皮層の最内層には、1層の細胞層からなる内皮 (endodermis) が存在する[9][15][31] (図2b)。内皮にはカスパリー線が存在し (細胞壁を通した物質輸送を遮断し、原形質を通した輸送のみを可能にしている)、中心柱への物質の出入りを調節している。古くなった内皮ではしばしばほとんどの細胞壁が木化し (中心柱からの水の漏出を防ぐ)、肥厚していない一部の内皮細胞 (通過細胞 passage cell) を通して通水する[15][16][24]。
内皮より内側の部分は、中心柱 (stele, central cylinder, central column) とよばれ、主に維管束からなる[15][23][32]。中心柱の周縁部には1〜数層の柔細胞からなる内鞘 (pericycle) があり、新たな側根はふつうここから (または内皮から) 生じる[15][23][24] (下記)。中心柱は放射中心柱 (actinostele) であり、中央に位置する木部 (一次木部) は横断面で放射状に突出部 (腕、ray) をもち、腕の間に師部 (一次師部) が位置する[9][15][24] (図2c)。木部の中心が髄になり、木部の腕がそれぞれ独立していることもある[15] (図2b)。木部はふつう外原型 (外側から求心的に形成される) であるが[9][15][23]、小葉植物の根では木部は内原型 (内側から遠心的に形成される) である[33][注釈 1]。
中心柱における木部の突出部 (腕) の数 (原生木部の数) は同一個体内でも変化することがあるが、ふつう種によってほぼ一定である[24]。根の木部は、原生木部の数に応じて二原型 (diarch)、三原型 (triarch; 図2c)、四原型 (tetrarch)、五原型 (pentarch) とよばれ、また6個以上の場合は多原型 (polyarch) とよばれる (単子葉類に多い; 図2a, b)[9][15][16][23]。ミズニラ属 (小葉植物) の根の中心柱は特異であり、一原型 (monoarch) である (古生代のリンボク類と共通)[23][35]。
分枝

根は、ふつう根端から比較的離れた場所で、側根 (lateral root; 分枝根 branch root[36]) を形成して側方分枝 (中軸分枝) する[14][12][15][16]。根の内部の中心柱の最外層にある内鞘 (またはその外側の内皮) から新たな側根の原基が生じ、これが皮層や表皮を突き破って伸長する (図3a)。すなわち根の分枝は内生的 (endogenous; 新たな根が内部に形成される) であり、茎の分枝が外生的 (exogenous; 新たな茎が表面から形成される) であるのとは対照的である[8][16]。
根はしばしば分枝を繰り返す。主となる根から生じた側根は一次側根 (primary lateral root)、そこから生じた側根は二次側根 (secondary lateral root) のように順によばれることがある[37]。
側根はふつう根の中心柱に対して特定の位置に由来し、特に原生木部に面する部分 (横断面で木部が外側へ突出している部分) から生じることが多いが、他にも原生師部に面する部分や原生木部と原生師部の間から生じる例も知られている[15][16]。そのため、側根は縦列 (または螺生) して生じることが多く[8]、その列数から中心柱の構造が推定できる。側根が2列であるダイコン (アブラナ科) は二原型木部、側根が4列であるニンジン (セリ科) は四原型木部、側根が5列であるサツマイモ (ヒルガオ科) は五原型木部をもつ[15]。

上記のように根の分枝はふつう内生的であり側方分枝であるが、例外的に小葉植物の根はその茎と同様に、根端分裂組織が2分することによって二又分枝する[38][39][40] (図3b)。つまり小葉植物の根の分枝は外生的 (新たな根が表面から形成される) である。また小葉植物は、根の木部が内原型である点でも他の維管束植物とは異なっている (上記)。このように小葉植物とそれ以外の維管束植物 (大葉植物、真葉植物) の根は大きく異なる特徴を示し、一般的にこれらの根は異なる起源をもつものと考えられている[38][39]。ただし小葉植物の根も、根冠や根毛をもつ点や、茎から内生発生する点では大葉植物の根と共通している。
根系
ある植物において地下部または根全体、あるいは1個の根とそこから生じている根を合わせたものは、根系 (こんけい、root system) とよばれる[41][8][42][3]。地下部全体とする場合、根系は根と共に地下茎なども含む[14]。この場合、維管束植物の植物体は、地上部のシュート系と地下部の根系からなる[43]。
太根と細根
太さに応じて根を太根と細根に類別することがある[37]。樹木では、一部の根が太く肥大し、それに細い根をまじえている。一方、イネ科の草本などでは、全ての根が肥大せず同様な太さになっている。このような中で、太く肥大した根を太根 (woody root, thick root)、太根を主とする根系は太根型根系 (woody root system) とよぶことがある。一方、細いままである根を細根 (fine root, rootlet)、細根を主とする根系は細根型根系 (fine root system) とよぶことがある。
定根と不定根



維管束植物において、根は胚の段階 (種子の中など) で幼根 (radicle) として形成される[10][37]。これが成長して一次根[44] (初生根[37]、primary root) となり、発達したものは主根[37][45][注釈 3] (main root、直根[10] taproot) になる。主根からは側根が生じる。このように幼根に由来する根、およびそこから生じた根を定根とよぶ[14]。定根からなる根系は、一次根系 (primary root system) とよばれ[14]、また主根型根系[37] (主根系[46]、直根系[3]、taproot system) ともよばれる (図4a)。
定根に対して、幼根以外に由来する根は不定根 (adventitious root) とよばれる[10][37][47]。不定根は、ふつう茎の維管束周辺から内生的に生じるが、まれに外生的に生じる例も知られている (例:ベゴニアの葉挿し)[16][48]。不定根は茎の節から生じることが多く、このような根は節根 (nodal root) ともよばれる[37] (図4c)。そのため、挿し木にはふつう節を残した茎が用いられる[24]。また定根と同様、不定根も側根を生じて側方分枝する (小葉植物以外; 上記参照)。シダ植物 (広義) や単子葉植物では、ふつうほとんどの根が不定根からなる。このような根系は二次根系 (secondary root system) または不定根系 (adventitious root system) とよばれる[14]。また多数まとまって生じている一様な不定根はひげ根 (fibrous root) とよばれ[37][49]、ひげ根からなる根系はひげ根型根系[37] (ひげ根系[3]、fibrous root system) とよばれる (図4b)。
種子植物において、種子から生じる根は種子根 (seminal root) とよばれる[10][37]。種子根はふつう幼根であるが、既に幼根から側根 (種子側根) が生じている例もある[50]。またイネ科などでは、胚軸から生じた不定根が種子根となることもある (種子不定根 seminal adventitious root)[50]。
一次成長と二次成長


上記のように、根は頂端分裂組織 (apical meristem) である根端分裂組織における細胞分裂とそれに続く細胞の拡大伸長によって成長する[4][19][20]。この成長は一次成長 (primary growth) とよばれ、基本的に長さを増す成長である[51]。一方、一次成長がほぼ完了した部位において新たな分裂組織が生じることがあり、これによる成長は二次成長 (secondary growth) とよばれる[51]。二次成長は基本的に太さを増す成長であり、これを司る分裂組織は側部分裂組織 (lateral meristem) である[51]。側部分列組織には、維管束形成層やコルク形成層がある。
単子葉植物などを除き、多くの種子植物の根は二次成長を行う[9][16][52]。一次木部と一次師部の間に (木部を取り囲むように) 維管束形成層 (vascular cambium) がつくられ、内側に二次木部を、外側に二次師部を形成していく[9][53]。根はふつう放射中心柱をもつため、維管束形成層の横断面は最初は星状だが、二次成長によってやがて円形になる[16][52][53]。木本植物の根では、茎と同様に二次木部は主に仮道管や道管要素、木部繊維など木化した細胞からなるが (図5a, b)、サツマイモ (ヒルガオ科) などの根では多量の柔細胞が形成される[54]。
茎の二次成長と同様、活発な二次成長によって直径が増すと、表皮、さらに皮層は裂けて剥がれることがある。この際には、皮層や内鞘などに1層の細胞からなる側部分裂組織であるコルク形成層 (phellogen) が形成される[9][24][55][56]。コルク形成層は外側にコルク組織 (phellem)、内側にコルク皮層 (phelloderm) を形成し、これらは合わせて周皮 (periderm) とよばれる。コルク組織の細胞は原形質を欠き、細胞壁にスベリン、ときにリグニンが沈着して根の表面を保護している (図5a, b)。
根の機能
根はふつう地中にあり、水やそこに含まれる窒素塩 (硝酸塩など) やカリウム、カルシウム、リン酸などの無機養分を吸収し、維管束の木部を通して植物体全体に送る (木部輸送)。根は分枝することで表面積を広げ、このような水や無機養分を吸収している。ライムギ (イネ科) の場合、根の表面積は地上部のシュート系 (茎と葉) の表面積の40倍に達すると試算されている[48]。根は効率的な無機栄養吸収のための応答を示し、例えば硝酸塩が多い場所では根はよく分枝し、またその細胞は効率よく硝酸塩を吸収できるような遺伝子発現を行う[57]。根毛や菌根菌の存在は根の表面積を広げ、根の吸収効率を高めている。
土壌粒子はふつう負に帯電しているため、硝酸、リン酸、硫酸などの陰イオンは土壌粒子には結合しない。そのためこれらの無機栄養は容易に土壌溶液に溶脱し、根によって吸収される[58]。一方、カリウム、カルシウム、マグネシウムなど陽イオンは土壌粒子に結合しており、容易には溶脱しない。根は呼吸によって土壌中に二酸化炭素を放出し、土壌溶液を酸性化する。その結果水素イオン (H+) が供給される。この水素イオンが土壌粒子を中和、結合していた陽イオンが土壌溶液に溶脱し、根が吸収する。この過程は陽イオン交換 (cation exchange) とよばれる[58]。
根の表面で吸収された無機養分を含む水溶液は、細胞壁内や細胞間隙など原形質外の通路 (アポプラスト経路) や原形質を通る通路 (シンプラスト経路) を通って維管束の木部へ輸送される[57]。根では、維管束は内皮に囲まれているため、吸収された水溶液が木部に輸送される際には必ず内皮を通る。内皮細胞どうしの接着部には疎水性物質であるスベリンが蓄積してカスパリー線が形成され、さらに細胞膜がカスパリー線に密着している[31][28][57]。そのため、アポプラスト経路で輸送されてきた水溶液も、内皮では細胞壁を通ることはできず、必ず内皮細胞の原形質を通らなければならない。内皮細胞は木部へ送られる物質の選別を行い、必要な物質を通し、不必要な物質は透過しない[57]。また、内皮細胞は中心柱から外側へ物質が逆流することを防いでいる[57]。さらに、皮層の最外層にカスパリー線をもつ外皮が形成されることもある (上記)。
根は植物ホルモンであるサイトカイニンの主な生成場所であり、他にもオーキシンやジベレリン、ストリゴラクトンなどの植物ホルモンを生成する[59]。サイトカイニンは細胞分裂を制御し、オーキシンは側根や不定根の形成を促進する[59]。またオーキシンは高濃度では細胞伸長を抑制するが、この伸長抑制が根の重力屈性に関わっていると考えられている[59]。エチレンによって根や根毛形成が促進され、ブラシノステロイドは低濃度で根の成長促進、高濃度で根の成長阻害をする[59]。ストリゴラクトンは菌根菌を根に誘因するが、ストリゴラクトンを感知して宿主の根に寄生する寄生植物も知られている[60]。
さまざまな根
根はふつう地中にあり、植物体の固定と水・無機養分の吸収という機能をもつ。しかし地中部にあってもこれ以外の機能をもつ根も存在する。また地中ではなく地上に伸びて機能する根もある (気根)。さらに、根はしばしば他生物 (菌根菌、根粒菌、宿主植物など) と密接な共生関係を結んでいる。
地中根
地中にある根は地中根 (terrestrial root) と総称される[37]。






- 貯蔵根 (storage root)[10][37][54]
- 肥大して水や養分を貯蔵する特殊化した根のこと。貯蔵された栄養は、一定期間の後に、または母体から離れて分散された後に新たな地上部を生じることに用いられる。以下では塊根と多肉根に類別したが[37]、その区分はかならずしも一定していない[61]。また球根とよばれるものは、塊根などのほかに、鱗茎や塊茎など貯蔵器官となった地下茎も含む。


- 収縮根 (contractile root; 牽引根 traction root)[10][16][37][62]
- クラスター根 (cluster root, proteoid root)[63]
- 短い側根が密生して試験管ブラシ状に変形した根 (図6h) であり、また有機酸分泌能力が一般的な根よりも高く、土壌中の難利用性のリンを溶解し吸収しやすくすることでリン欠乏土壌に適応している。ヤマモガシ科 (学名: Proteaceae) の植物から発見されたため、かつては proteoid root とよばれていた。しかし後にマメ科、クワ科、ヤマモモ科などからも見つかったため、形態的特徴に基づいてクラスター根(房のような根の意味)とよばれるようになった。また側根ではなく根毛が房状に形成されたダウシフォーム根(dauciform root)がカヤツリグサ科やイグサ科の一部に、同様のキャピラロイド根がサンアソウ科に見られ、これらもクラスター根と同様にリン吸収に適応したものであると考えられている[64]。
気根
地上部にある根は気根 (aerial root) と総称される[37][65]。地下茎から生じるものや、地上茎、水中茎から生じるものなどがある。





- 板根 (buttress root)[37][69]
- 支柱根 (支持根、prop root、支柱気根、支持気根、prop aerial root)[10][37][65] (→詳細は「支柱根」を参照)




- 付着根 (着生根[54][71]、adhesive root, adhering root、よじのぼり根[72]、climbing root)[10][37][65]
- 根性巻ひげ (root tendril)[54]


水中根

通常の状態として水中に伸びている根を水中根 (aquatic root) という[37] (図8)。このような根は、根冠や根毛を欠いていることがある[8][26]。ミズキンバイ (アカバナ科) は、水底を横走する根茎の背面から列状に生じて水中に浮かんでいる根をもち、特に浮根 (floating root) とよばれる[37]。
他生物と共生した根
根はふつうは地中にあり、他生物と密接な共生関係を築いている例が多い。根は特に根冠や根毛を通じて有機物 (光合成産物の20%にも達することもある) を土壌中に分泌・放出しており、根の周囲に特異な環境を形成している[58]。このような環境は根圏 (rhizosphere) とよばれ、さまざまな微生物が植物と共生関係を結んで生育している。また下記のように、ほとんどの維管束植物は根において菌類と直接的に共生して菌根を形成しており、さらに窒素固定を行う生物と共生して特異な構造を形成している例もある。


- 菌根 (mycorrhiza, pl. mycorrhizae)[37] (→詳細は「菌根」を参照)
- 維管束植物のほとんどは根において菌類 (菌根菌、mycorrhizal fungus) と共生し、菌根を形成している[74]。ただし水生植物やウラボシ科、アブラナ科、ヒユ科、ナデシコ科、タデ科などでは菌根をもたない種が比較的多く知られている。菌根の形態や菌根菌のグループにはさまざまなタイプが知られており、それに応じて外生菌根 (外菌根; 図9a, 9c-c)、アーバスキュラー菌根 (図9b, 9c-b)、ツツジ型菌根 (エリコイド菌根)、イチヤクソウ型菌根 (アルブトイド菌根)、シャクジョウソウ型菌根 (モノトロポイド菌根)、ラン型菌根 (図9c-a) などに類別されている[75]。この中ではアーバスキュラー菌根が最も普遍的であり、進化的にも最も祖先的な菌根であると考えられている[74]。根が合成する植物ホルモンであるストリゴラクトンは、アーバスキュラー菌根菌を根に誘引する[59]。菌根菌が根の表層や細胞間隙に菌糸を張り巡らせるものや、植物細胞内 (正確には細胞壁と細胞膜の間) に侵入して栄養交換用の構造を形成するものがいる[58][76] (図9c)。菌根菌の菌糸は根毛よりも細く、遥かに長く土壌中に張巡らされており、より効率的に無機養分や水を吸収し、これを植物に供給している[77]。また菌根菌は、植物に病害や乾燥ストレスに対する耐性を付与することも知られている[78]。一方、植物は菌根菌に有機物を与えており、菌根菌との間に相利共生関係が築かれている。ただし植物の中には、自らは光合成せずに有機物も菌根菌から得ている例がある (腐生植物 = 菌従属栄養植物、菌寄生植物)[79]。また菌根菌は、異種間を含むさまざまな植物の根をつなぎ (菌根菌ネットワーク)、その間で糖などの物質転送が起こっていることが知られている[76][80]。


- 根粒 (根瘤、root nodule)[37] (→詳細は「根粒」を参照)
- マメ科の植物では、根に根粒菌と総称される窒素固定能をもつ細菌が共生し、根粒とよばれる粒状の構造を形成する (図9d, e)。根粒菌は窒素化合物を供給し、植物は有機物を供給する相利共生関係が築かれている。マメ科植物と共生する根粒菌はプロテオバクテリア門に属するが、マメ目に比較的近縁なバラ目 (グミ)、ブナ目 (ヤマモモ、ハンノキ、モクマオウ)、ウリ目 (ドクウツギ、ナギナタソウ) の中には、窒素固定能をもつ放線菌のフランキア属と共生して根粒を形成するものが知られている[81]。このような植物はアクチノリザル植物 (actinorhizal plant)、形成される根粒は放線菌根 (actinorhiza) やハンノキ型根粒ともよばれる[82][83] (図9f)。マメ目、バラ目、ブナ目、ウリ目は単系統群を形成しており、この系統群は窒素固定クレードとよばれる[84]。根粒形成の機構は、アーバスキュラー菌根形成の機構をもとにしたものであることが示されている[84]。


寄生根


共生の1形態として、寄生がある。他の植物に寄生し養分を奪う植物は寄生植物とよばれ、自ら光合成を行いながら宿主からも栄養分を奪う半寄生植物 (ヤドリギなど) と、光合成能を欠き、有機物も含めた栄養分を宿主から奪う全寄生植物 (ネナシカズラなど) がある[88]。寄生植物は栄養分を吸収するために宿主に吸器 (haustorium, pl. haustoria) を付着させているが、寄生植物における吸器は特殊化した根であり、この根は寄生根 (parasitic root) ともよばれる[10][16][37]。寄生根では、しばしば寄生植物と宿主の維管束 (木部) がつながっている (木部架橋、xylem bridge)[24] (図10b)。寄生根は、以下のようにいくつかのタイプに類別されることがある[37][54]。
- シオガマギク型 (Pedicularis type)
- ハマウツボ型 (Orobanche type)
- ヤドリギ型 (Viscum type) (図10a)
- ネナシカズラ型 (Cuscuta type) (図10b)
人間との関わり


根菜とよばれる野菜の中には、サトイモ (サトイモ科)、タマネギ (ヒガンバナ科)、レンコン (ハス科)、ジャガイモ (ナス科) など実際には根ではなく茎 (根茎、塊茎など) に由来するものも多い。根 (ときにそれに続く胚軸も含めて) を食用として利用するものとしては、ダイコンやカブ、ハツカダイコン、ホースラディッシュ、ルタバガ、マカ (アブラナ科)、キャッサバ (トウダイグサ科)、クズ、ホドイモ (マメ科)、ビート、テンサイ (図11a) (ヒユ科)、サツマイモ (図11b) (ヒルガオ科)、ニンジン、パースニップ (セリ科)、ゴボウ、モリアザミ、サルシファイ、ヤーコン (キク科) などがある[10][37][89][90][91][92]。


一方、薬用とされる根もあり (地下茎と区別せずに共に用いられる例もある)、テンダイウヤク (クスノキ科)、ジャノヒゲ (キジカクシ科)、トリカブト、サキシマボタンヅル (キンポウゲ科)、シャクヤク、ボタン (ボタン科)、キバナオウギ、カンゾウ、クララ (マメ科)、イトヒメハギ、セネガ (ヒメハギ科)、オオカラスウリ (ウリ科)、ヒナタイノコヅチ (ヒユ科)、ツルドクダミ (タデ科)、トコン (アカネ科)、ゲンチアナ (図11c)、トウリンドウ (リンドウ科)、インドジャボク (キョウチクトウ科)、ムラサキ (ムラサキ科)、コガネバナ (シソ科)、ベラドンナ、ハシリドコロ (ナス科)、オタネニンジン (高麗人蔘; 図11d) (ウコギ科)、ミシマサイコ、ノダケ、トウキ、トウスケボウフウ、ヨロイグサ (セリ科)、キキョウ (キキョウ科)、カノコソウ、オミナエシ (スイカズラ科)、モッコウ (キク科) の根が利用される[93]。
またアカネ (アカネ科) や ムラサキ (ムラサキ科) の根は、古くから染料として用いられてきた[94]。


上記のように、マメ科植物の多くは根において窒素固定細菌と共生して根粒を形成している。そのため、耕作地にマメ科植物 (シロツメクサ、ミヤコグサなど) を栽培し、窒素栄養分などを土地に供給する緑肥として利用することがある (図11e)。マメ科植物の利用は、18世紀の農業革命において重要な役割を演じた[95]。
根は地中を伸長し、また肥大成長することで母岩などを破壊し、このような働きは土壌形成に重要な役割を果たしている。このような働きにより、舗装道路など人工構造が破壊されることもある。また根の成長によって、アンコール遺跡などの遺跡が被害を受けることもある (一方でこのような景観が観光スポットにもなっている)[96] (図11f)。
脚注
注釈
出典
- ^ 農業用語の基礎知識:営農・園芸のすべてがわかる必携用語集 著者: 藤重宣昭 p.310
- ^ 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “根”. 岩波 生物学辞典 第5版. 岩波書店. p. 1053. ISBN 978-4000803144
- ^ a b c d 原襄 (1994). “植物の基本構造”. 植物形態学. 朝倉書店. pp. 5–11. ISBN 978-4254170863
- ^ a b c d e f g h i j k l 池内昌彦, 伊藤元己, 箸本春樹 & 道上達男 (監訳) (2018). “35 維管束植物の構造、生長、発生”. キャンベル生物学 原書11版. 丸善出版. pp. 869–897. ISBN 978-4621302767
- ^ アーネスト・ギフォード & エイドリアンス・フォスター (著) 長谷部光泰, 鈴木武 & 植田邦彦 (監訳) (2002). “シュートと根”. 維管束植物の形態と進化. 文一総合出版. pp. 32–33. ISBN 978-4829921609
- ^ 加藤雅啓 (編) (1997). “2-1 マツバラン綱”. バイオディバーシティ・シリーズ (2) 植物の多様性と系統. 裳華房. pp. 198–199. ISBN 978-4-7853-5825-9
- ^ アーネスト・ギフォード & エイドリアンス・フォスター (著) 長谷部光泰, 鈴木武 & 植田邦彦 (監訳) (2002). “サンショウモ目”. 維管束植物の形態と進化. 文一総合出版. pp. 321–325. ISBN 978-4829921609
- ^ a b c d e 熊沢正夫 (1979). “根の通性”. 植物器官学. 裳華房. pp. 304−312. ISBN 978-4785358068
- ^ a b c d e f g h i j k 河野恭廣 (1998). “根の基本的構造”. In 根の事典編集委員会 (編). 根の事典. 朝倉書店. pp. 1–2. ISBN 978-4254420210
- ^ a b c d e f g h i j k l m n o 原襄 (1994). “根”. 植物形態学. 朝倉書店. pp. 12–16. ISBN 978-4254170863
- ^ 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “根端”. 岩波 生物学辞典 第5版. 岩波書店. pp. 502–503. ISBN 978-4000803144
- ^ a b c d 原襄 (1994). “根端”. 植物形態学. 朝倉書店. pp. 129–134. ISBN 978-4254170863
- ^ a b 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “根端分裂組織”. 岩波 生物学辞典 第5版. 岩波書店. p. 503. ISBN 978-4000803144
- ^ a b c d e f g h i 清水建美 (2001). “根”. 図説 植物用語事典. 八坂書房. pp. 233–236. ISBN 978-4896944792
- ^ a b c d e f g h i j k l m n o p q r s t u v w 原襄 (1994). “根の構造”. 植物形態学. 朝倉書店. pp. 47–55. ISBN 978-4254170863
- ^ a b c d e f g h i j k l m ポーラ・ルダル (著) 鈴木三男 & 田川裕美 (翻訳) (1997). “茎の肥大成長”. 植物解剖学入門 ―植物体の構造とその形成―. 八坂書房. pp. 55–69. ISBN 978-4896946963
- ^ a b 飯嶋盛雄 (1998). “根冠”. In 根の事典編集委員会 (編). 根の事典. 朝倉書店. pp. 3–5. ISBN 978-4254420210
- ^ 池内昌彦, 伊藤元己, 箸本春樹 & 道上達男 (監訳) (2018). “重力”. キャンベル生物学 原書11版. 丸善出版. p. 983. ISBN 978-4621302767
- ^ a b 森田茂紀 (1998). “根端分裂組織”. In 根の事典編集委員会 (編). 根の事典. 朝倉書店. pp. 5–6. ISBN 978-4254420210
- ^ a b 谷本英一 (1998). “根の生長と細胞分裂”. In 根の事典編集委員会 (編). 根の事典. 朝倉書店. pp. 30–33. ISBN 978-4254420210
- ^ a b c L. テイツ, E. ザイガー, I.M. モーラー & A. マーフィー (編) (2017). “根の成長と分化”. 植物生理学・発生学 原著第6版. 講談社. pp. 544–549. ISBN 978-4061538962
- ^ 河合義隆 (1998). “双子葉植物における根の始原体の形成”. In 根の事典編集委員会 (編). 根の事典. 朝倉書店. pp. 24–26. ISBN 9784254420210
- ^ a b c d e f g h i j 清水建美 (2001). “根の内部構造”. 図説 植物用語事典. 八坂書房. pp. 246–249. ISBN 978-4896944792
- ^ a b c d e f g h i j Rudall, P. J. (2007). “Root”. Anatomy of Flowering Plants: An Introduction to Structure and Development. Cambridge University Press. pp. 43–56. ISBN 978-0521692458
- ^ 原襄 (1994). “表皮”. 植物形態学. 朝倉書店. pp. 75–76. ISBN 978-4254170863
- ^ a b c d e 森田茂紀 (1998). “根毛”. In 根の事典編集委員会 (編). 根の事典. 朝倉書店. pp. 7–8. ISBN 978-4254420210
- ^ 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “根毛”. 岩波 生物学辞典 第5版. 岩波書店. p. 506. ISBN 978-4000803144
- ^ a b 森田茂紀 (1998). “皮層”. In 根の事典編集委員会 (編). 根の事典. 朝倉書店. pp. 8–10. ISBN 978-4254420210
- ^ 巽二郎 (1998). “通気”. In 根の事典編集委員会 (編). 根の事典. 朝倉書店. pp. 14–16. ISBN 978-4254420210
- ^ 山内章 (1998). “下皮”. In 根の事典編集委員会 (編). 根の事典. 朝倉書店. pp. 10–12. ISBN 978-4254420210
- ^ a b 唐原一郎 (1998). “内皮とカスパリー線”. In 根の事典編集委員会 (編). 根の事典. 朝倉書店. pp. 12–14. ISBN 978-4254420210
- ^ Beck, C. B. (2005). “Primary tissues and tissue regions”. An Introduction to Plant Structure and Development. Cambridge University Press. pp. 281–289. ISBN 978-0521837408
- ^ Simpson, M. (2006). Plant Systematics. Academic Press. p. 78. ISBN 978-0126444605
- ^ 長谷部光泰 (2020). 陸上植物の形態と進化. 裳華房. p. 132. ISBN 978-4785358716
- ^ アーネスト・ギフォード & エイドリアンス・フォスター (著) 長谷部光泰, 鈴木武 & 植田邦彦 (監訳) (2002). 維管束植物の形態と進化. 文一総合出版. pp. 168–169. ISBN 978-4829921609
- ^ 山内章 (1998). “根の分枝”. In 根の事典編集委員会 (編). 根の事典. 朝倉書店. pp. 33–35. ISBN 978-4254420210
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj 清水建美 (2001). “根の分類”. 図説 植物用語事典. 八坂書房. pp. 236–246. ISBN 978-4896944792
- ^ a b 加藤雅啓 (編) (1997). “7-2-3 根”. バイオディバーシティ・シリーズ (2) 植物の多様性と系統. 裳華房. pp. 83–84. ISBN 978-4-7853-5825-9
- ^ a b 加藤雅啓 (1998). “原始的維管束植物の体制と根の起源”. In 根の事典編集委員会 (編). 根の事典. 朝倉書店. pp. 152–154. ISBN 978-4254420210
- ^ アーネスト・ギフォード & エイドリアンス・フォスター (著) 長谷部光泰, 鈴木武 & 植田邦彦 (監訳) (2002). “ヒカゲノカズラ属 器官学”. 維管束植物の形態と進化. 文一総合出版. pp. 116–119. ISBN 978-4829921609
- ^ 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “根系”. 岩波 生物学辞典 第5版. 岩波書店. p. 501. ISBN 978-4000803144
- ^ 中元朋実 (1998). “根系の形態”. In 根の事典編集委員会 (編). 根の事典. 朝倉書店. p. 76. ISBN 978-4254420210
- ^ 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “シュート系”. 岩波 生物学辞典 第5版. 岩波書店. pp. 643–644. ISBN 978-4000803144
- ^ 日本植物学会 (1990). 文部省 学術用語集 植物学編 (増訂版). 丸善. p. 533. ISBN 978-4621035344
- ^ a b 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “主根”. 岩波 生物学辞典 第5版. 岩波書店. p. 633. ISBN 978-4000803144
- ^ 日本植物学会 (1990). 文部省 学術用語集 植物学編 (増訂版). 丸善. p. 587. ISBN 978-4621035344
- ^ 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “不定根”. 岩波 生物学辞典 第5版. 岩波書店. p. 1207. ISBN 978-4000803144
- ^ a b Bowes, B. & Mauseth, J. D. (2008). “The Root”. Plant Structure: A Colour Guide 2nd Edition. Jones & Bartlett Learning. pp. 185–189. ISBN 978-0763763862
- ^ 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). 岩波 生物学辞典 第5版. 岩波書店. p. 1139. ISBN 978-4000803144
- ^ a b 山内章 (1993). “根の種類 (1)”. 根の研究 2: 20-23. doi:10.3117/rootres.2.20.
- ^ a b c 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “分裂組織”. 岩波 生物学辞典 第5版. 岩波書店. p. 1256. ISBN 978-4000803144
- ^ a b 田中典幸 (1998). “根の2次肥大生長”. In 根の事典編集委員会 (編). 根の事典. 朝倉書店. pp. 36–37. ISBN 978-4254420210
- ^ a b 原襄 (1994). “形成層と二次組織”. 植物形態学. 朝倉書店. pp. 134–139. ISBN 978-4254170863
- ^ a b c d e f g 熊沢正夫 (1979). “異形根”. 植物器官学. 裳華房. pp. 313−325. ISBN 978-4785358068
- ^ 原襄 (1994). “コルク形成層と周皮”. 植物形態学. 朝倉書店. pp. 139–141. ISBN 978-4254170863
- ^ 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “コルク形成層”. 岩波 生物学辞典 第5版. 岩波書店. p. 493. ISBN 978-4000803144
- ^ a b c d e 池内昌彦, 伊藤元己, 箸本春樹 & 道上達男 (監訳) (2018). “36 維管束植物の栄養吸収と輸送”. キャンベル生物学 原書11版. 丸善出版. pp. 899–920. ISBN 978-4621302767
- ^ a b c d 池内昌彦, 伊藤元己, 箸本春樹 & 道上達男 (監訳) (2018). “37 土壌と植物の栄養”. キャンベル生物学 原書11版. 丸善出版. pp. 921–939. ISBN 978-4621302767
- ^ a b c d e 池内昌彦, 伊藤元己, 箸本春樹 & 道上達男 (監訳) (2018). “39 内外のシグナルに対する植物の応答”. キャンベル生物学 原書11版. 丸善出版. pp. 963–994. ISBN 978-4621302767
- ^ 米山弘一, 謝肖男 & 米山香織 (2010). “根寄生植物の発芽シグナルとしてのストリゴラクトン”. 植物の生長調節 45: 83-94. doi:10.18978/jscrp.45.2_83.
- ^ a b 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “塊根”. 岩波 生物学辞典 第5版. 岩波書店. p. 180. ISBN 978-4000803144
- ^ 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “収縮根”. 岩波 生物学辞典 第5版. 岩波書店. p. 623. ISBN 978-4000803144
- ^ de Kroon, H. & Visser, E.J.W. (著) 森田茂紀 & 田島亮介 (監修) (2008). 根の生態学. シュプリンガー・ジャパン. p. 22. ISBN 978-4431727354
- ^ 丸山隼人・和崎淳 (2017). “低リン条件で房状の根を形成する植物の機能と分布 -低リンストレスに対する植物の適応機構-”. 化学と生物 55 (3): 189-195. doi:10.1271/kagakutoseibutsu.55.189.
- ^ a b c 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “気根”. 岩波 生物学辞典 第5版. 岩波書店. pp. 283–284. ISBN 978-4000803144
- ^ 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “呼吸根”. 岩波 生物学辞典 第5版. 岩波書店. p. 471. ISBN 978-4000803144
- ^ 札幌市公園緑化協会 豊平公園緑のセンター (2012). 札幌市 緑のセンターだより 156.
- ^ 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “同化根”. 岩波 生物学辞典 第5版. 岩波書店. p. 977. ISBN 978-4000803144
- ^ 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “板根”. 岩波 生物学辞典 第5版. 岩波書店. p. 1119. ISBN 978-4000803144
- ^ 多田多恵子 (2002). したたかな植物たち―あの手この手のマル秘大作戦. エスシーシー. pp. 126–133. ISBN 978-4886479228
- ^ 日本植物学会 (1990). 文部省 学術用語集 植物学編 (増訂版). 丸善. p. 320. ISBN 978-4621035344
- ^ 日本植物学会 (1990). 文部省 学術用語集 植物学編 (増訂版). 丸善. p. 366. ISBN 978-4621035344
- ^ 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “同化根”. 岩波 生物学辞典 第5版. 岩波書店. p. 502. ISBN 978-4000803144
- ^ a b Wang, B. & Qiu, Y. L. (2006). “Phylogenetic distribution and evolution of mycorrhizas in land plants”. Mycorrhiza 16: 299-363. doi:10.1007/s00572-005-0033-6.
- ^ 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “菌根”. 岩波 生物学辞典 第5版. 岩波書店. pp. 333–335. ISBN 978-4000803144
- ^ a b 池内昌彦, 伊藤元己, 箸本春樹 & 道上達男 (監訳) (2018). “31 菌類”. キャンベル生物学 原書11版. 丸善出版. pp. 753–773. ISBN 978-4621302767
- ^ 市石博 (2007). “学校便り(3)生態系をみる新たな視点 土の中に広がるネットワーク『菌根菌』研究の現場を見聞きして”. 日本生態学会誌 57: 277–280. doi:10.18960/seitai.57.2_277.
- ^ 松崎克彦 (2009). “アーバスキュラー菌根菌とその利用”. 農業および園芸 841: 170-175.
- ^ 辻田有紀 & 遊川知久 (編) (2014). “光合成をやめた植物ー菌従属栄養植物のたどった進化の道のり”. 植物科学最前線 5: 84–139.
- ^ 宝月岱造 (2010). “外生菌根菌ネットワークの構造と機能”. 土と微生物 64: 57–63. doi:10.18946/jssm.64.2_57.
- ^ 山中高史 & 岡部宏秋 (2008). “わが国に生育する放線菌根性植物とフランキア菌”. 森林総合研究所研究報告 7: 67–80. NAID 40016000067.
- ^ 九町健一 (2013). “共生窒素固定放線菌フランキア”. 生物工学会誌 91: 24-27. NAID 110009580287.
- ^ 植村誠次 (1977). “根粒菌と根粒植物”. URBAN KUBOTA 14: 22–25.
- ^ a b 林誠 (2015). “植物の窒素固定:植物と窒素固定細菌との共生の進化”. 領域融合レビュー 4: e010. doi:10.7875/leading.author.4.e010.
- ^ アーネスト・ギフォード & エイドリアンス・フォスター (著) 長谷部光泰, 鈴木武 & 植田邦彦 (監訳) (2002). “背地性根”. 維管束植物の形態と進化. 文一総合出版. p. 370. ISBN 978-4829921609
- ^ 加藤雅啓 (編) (1997). “3-3 ソテツ綱”. バイオディバーシティ・シリーズ (2) 植物の多様性と系統. 裳華房. pp. 218–219. ISBN 978-4-7853-5825-9
- ^ Cox, P.A., Banack, S.A. & Murch, S.J. (2003). “Biomagnification of cyanobacterial neurotoxins and neurodegenerative disease among the Chamorro people of Guam”. Proc. Natl. Acad. Sci. U.S.A. 100: 13380-13383. doi:10.1073/pnas.2235808100.
- ^ 清水建美 (2001). “有機栄養に関する区分”. 図説 植物用語事典. 八坂書房. pp. 13–15. ISBN 978-4896944792
- ^ 牧野晩成 (1978). 果物と野菜の観察. ニュー・サイエンス社. pp. 58–70. ASIN B000J8B2FA
- ^ 牧田道夫 (1994). “我が国が未利用の資源植物に関する調査”. 農業生物資源研究所研究資料 6: 103-172.
- ^ 日本食品標準成分表2015年版(七訂). 文部科学省.
- ^ 藤本滋生 (1984). “「葛粉 (くづこ) 一覧」 および 「澱粉 (くずこ) 一覧」 について”. 鹿兒島大學農學部學術報告 34: 17-28.
- ^ 薬用植物総合情報データベース. 薬用植物資源研究センター. (2020年5月27日閲覧)
- ^ 下山進, 下山裕子 & 大下浩司 (2017). “衣裳を彩る色材の分析―日本における染色の歴史と琉球紅型衣装にみられる色材―”. 文化財情報学研究: 吉備国際大学文化財総合研究センター紀要 14: 53-62. NAID 40021343738.
- ^ 間藤徹 (2015). “有機農業 2.0”. 日本農薬学会誌 40: 31-34. doi:10.1584/jpestics.W14-39.
- ^ 古部浩 (2004). “カンボジアのアンコール遺跡とその修復”. 地学雑誌 113: 539-544. doi:10.5026/jgeography.113.4_539.
関連項目
- 構造:根端分裂組織、根冠、根毛
- 根のいろいろ:呼吸根、支柱根
- 共生:菌根、菌根菌、根粒、根圏
- 園芸:球根、直根性、根詰まり、接ぎ木(根側に問題のある植物を問題のない木の根に接ぐ。)
- 土壌/ 腐植物質 / 土壌改良 / 水耕栽培
- 根圧 ‐ 浸透圧などを利用して根が吸い上げた水によって、内部の道管内の液体を輸送する。
- 通気組織 ‐ 水草などにある根や茎などにある空気を通す組織。
- 付着器(仮根) ‐ 海藻が海底に生やして流されないようにするもの。
外部リンク
