Хаплогрупа N-M231
| Хаплогрупа N | |
|---|---|
 | |
| Можно време на потекло | 36,800 [95% CI 34,300–39,300] п.н.е.[1] 44,700 или 38,300 п.н.е. во зависност од стапката на мутација[2] 41,900 [95% CI 40,175-43,591] п.н.е.[3] |
| Возраст на спојување | 21,700 [95% CI 19,500–23,900] п.н.е.[1] 25,313 [95% CI 21,722–28,956] п.н.е.[3] |
| Можно место на потекло | Северна Источна Азија[4][5] |
| Предок | NO |
| Својствени мутации | M231 |
Хаплогрупата N (M231) — хаплогрупа на ДНК на Y-хромозомот дефинирана со присуството на маркерот M231 на еднонуклеотиден полиморфизам (SNP).
Најчесто се среќава кај мажјаците со потекло од северна Евроазија. Исто така, забележано е на пониски честоти кај популациите родени во други региони, вклучувајќи делови од Балканот, Средна Азија, Источна Азија и Југоисточна Азија.
Потекло
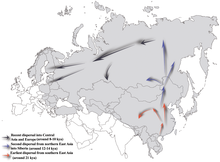
Хаплогрупата NO-M214 – нејзиниот најнов заеднички предок со својот брат или сестра, хаплогрупата O-M175 – се проценува дека постоела пред околу 36.800–44.700 години.[1][7]
Генерално се смета дека N-M231 се појавил во Источна Азија пред приближно 19.400 (±4.800) години и ја населил северна Евроазија по последниот глацијален максимум. Мажјаците кои го носеа маркерот очигледно се движеа кон север како што климата се загреваше во холоценот, мигрирајќи се во спротивна насока од стрелките на часовникот, за на крајот да се концентрираат во подалечните области како Феноскандија и Балтикот. Очигледниот недостаток на хаплогрупата N-M231 кај домородните американски народи покажува дека таа се проширила откако Берингија била потопена, пред околу 11.000 години.
Дистрибуција

Хаплогрупата N има широка географска дистрибуција низ северна Евроазија, а повремено е забележана и во други области, вклучително и Средна Азија и Балканот.
Најчест е кај домородните народи на Русија, вклучувајќи ги и финските народи, Мари, Удмурт, Коми, Ханти, Манси, Ненец, Нганасани, турски народи (јакути, Долгани, Хакаси, Тувани, Татари, Чуваши итн.). Бурјати, Тунгуски народи (Евенки, Евни, Негидали, Нанаи, итн.), Јукагири, Луораветланци (Чукчи, Корјаци) и сибирски ескими, но одредени подклади се многу чести во Финска, Естонија, Летонија и други подклади во Литванија. најдени на ниска честота во Кина (Ји, Накси, Лоба, Хан Кинези, итн.). Особено кај етничките фински народи и народите што зборуваат балтичко од северна Европа, народите кои зборуваат обугрички и северните самојед од западен Сибир и народите што зборуваат турски јазик во Русија (особено Јакутите, но исто така и Алтајци, Шорци, Каки, Чуваши, Татари и Башкири). Скоро сите членови на хаплогрупата N меѓу овие популации на северна Евроазија припаѓаат на подкладите или на хаплогрупата N-Tat или на хаплогрупата N-P43.
Y-хромозомите кои припаѓаат на N1b-F2930/M1881/V3743 или N1*-CTS11499/L735/M2291 (xN1a-F1206/M2013/S11466), се пронајдени во Кина и спорадично низ другите делови на Евроазија. N1a-F1206/M2013/S11466 се наоѓа во голем број во Северна Евроазија.
N2-Y6503, другата примарна подклада на хаплогрупата N, е исклучително ретка и е главно претставена меѓу постоечките луѓе со неодамна формирана подклада која е практично ограничена на земјите што ја сочинуваат поранешна Југославија (Босна и Херцеговина, Хрватска, Србија и Црна Гора), Унгарија и Австрија. Други членови на N2-Y6503 вклучуваат Унгарец со неодамнешно потекло од Сучеава во Буковина, Словак, неколку Британци и Алтајец.[1]
N* (M231)
За Y-хромозомите што ја прикажуваат мутацијата M231 што ја дефинира Хаплогрупата N-M231, но не ги прикажуваат мутациите CTS11499, L735, M2291 што ја дефинираат Хаплогрупата N1, се вели дека припаѓаат на парагрупата N-M231*.[4]
N-M231* е пронајден на ниски нивоа во Кина.[4] Од примерок од 165 мажи Хан од Кина, откриено е дека две лица (1,2%) припаѓаат на N*. Еден потекнува од Гуангжу и еден од Ксиан.
Меѓу античките примероци од бајкалската ранонеолитска култура Китои, еден од примероците Шаманка II (DA250), датиран во в. 6500 BP, беше анализиран како NO1-M214 во оригиналната студија.[8] Сепак, овој ист примерок (DA250 или Шаманка 250) подоцна беше откриено дека припаѓа на N-FT210118, истиот клад како и другите примероци од хаплогрупата N од истата локација (покрај DA247, кој припаѓа на N-Y147969). N-FT210118 е изведен од N-L666/N-F2199, но базален на N-CTS6380, ова е најновиот заеднички предок на денешниот N-P43 (се наоѓа главно кај Марис, Удмурт, Коми, Чуваши, Татари, Ненец, Нганасани, Канти, Манси, Кака, Туванци, итн.) и N-F1101 (се среќава главно кај источните Азијци). Понатаму, N-FT210118 не е пронајден кај ниту една жива личност која до денес ја тестирала неговата Y-DNA, а проценетиот TMRCA на N-CTS6380 го надминува проценетиот датум на таложење на кој било од примероците од локацијата Шаманка поврзана со Културата на Китои, па се чини дека претставниците на културата Китои во Шаманка (или барем нивната Y-DNA) изумреле наместо да бидат директни предци на кој било жив народ.[9][10]
N1 (CTS11499, Z4762, CTS3750)
Во 2014 година, имаше голема промена во дефиницијата за подкладата N1, кога LLY22g беше повлечен како главен дефинирачки SNP за N1 поради извештаите за несигурноста на LLY22g. Според ISOGG, LLY22g е проблематичен бидејќи е „палиндромски маркер и лесно може погрешно да се протолкува“. Оттогаш, името N1 се применува на клад означен со голем број SNP, вклучувајќи ги CTS11499, Z4762 и CTS3750. N1 е најновиот заеднички предок на сите постоечки членови на Хаплогрупата N-M231 освен членовите на ретката N2-Y6503 (N2-B482) подклада. TMRCA на N1 се проценува дека е 18.000 п.н.е. (16.300-19.700 BP; 95% CI).[1] Од ревизијата во 2014 година, позицијата на многу примери на „N1-LLY22g“ во хаплогрупата N стана нејасна.
Сепак, во постарите студии, N-LLY22g е пријавено дека достигнува честота до 30% (13/43) кај луѓето Ји од округот Бутуо, Сечуан во Југозападна Кина (Hammer и др. 2005, Karafet и др. 2001 и Wen и др. 2004). Тоа е исто така пронајдено кај 34,6% од луѓето Лхоба (Wen и др. 2004).[11] N1-LLY22g* е пронајден во примероци од хански кинески, но со широко различна честота:
- 6,8% (3/44) Хан од Ксиан (Hammer и др. 2005 и Karafet и др. 2001)
- 6,7% (2/30) Хан од Ланжу Xue и др. 2006
- 3,6% (3/84) тајвански Хан Hammer и др. 2005
- 2,9% (1/34) Хан од Ченгду Xue и др. 2006
- 2,9% (1/35) Хан од Харбин Xue и др. 2006
- 2,9% (1/35) Хан од округот Меиксиан Xue и др. 2006
- 0% (0/32) Хан од градот Јининг Xue и др. 2006
Други популации во кои се пронајдени претставници на N1*-LLY22g вклучуваат:
- Луѓето од Хани (4/34 = 11,8%) Xue и др. 2006
- Сибе луѓе (4/41 = 9,8%) Xue и др. 2006
- Луѓе Туџија (2/49 = 4,1%) Hammer и др. 2005
- Луѓе од Манчу (2/52 = 3,8% Hammer и др. 2005 до 2/35 = 5,7% Xue и др. 2006
- Бит луѓе (1/28 = 3,6%) Cai 2011
- Ујгури (2/70 = 2,9% Xue и др. 2006 до 2/67 = 3,0%) Hammer и др. 2005
- Тибетски народ (3/105 = 2,9% Hammer и др. 2005 до 3/35 = 8,6% Xue и др. 2006)
- Корејци (0/106 = 0,0% - 2/25 = 8% (Rootsi и др. 2006, Xue и др. 2006, и Kim 2007)
- Виетнамци (2/70 = 2,9%) Hammer и др. 2005
- Јапонци (0/70 Токушима – 2/26 = 7,7% Аомори) Hammer и др. 2005
- Manchurian Evenks (0/26 = 0,0% Xue и др. 2006 до 1/41 = 2,4% Hammer и др. 2005)
- Луѓето од Алтај (0/50 северни до 5/96 = 5,2% јужни, или 0/43 Бешпелтир до 5/46 = 10,9% Кулада), Hammer и др. 2005 Kharkov 2007
- Шорс (2/23 = 8,7%) Rootsi и др. 2006
- Луѓето од Кака (5/181 = 2,8%) Rootsi и др. 2006
- Tuvans (5/311 = 1,6%) Rootsi и др. 2006
- Јужен Борнео (1/40 = 2,5%) Rootsi и др. 2006
- Forest Nenets (1/89 = 1,1%) Rootsi и др. 2006
- Јакути (0/215 - 1/121 = 0,8%) Rootsi и др. 2006
- Турски народ (1/523 = 0,2%) Rootsi и др. 2006; Во Турција, вкупниот број на подклади на хаплогрупата N-M231 изнесува 4% од машката популација.
- Еден поединец кој припаѓа или на N* или N1* е пронајден во примерок од 77 мажи од Катманду, Непал (1/77 = 1,3% N-M231(xM128,P43,Tat)) Gayden 2007
N1(xN1a,N1c) е пронајден во античките коски на цивилизацијата Лиао:[12]
- Ниухелијанг (Хонгшанска култура, 6500–5000 БП) 66,7% (=4/6)
- Халахаигу (Култура на Ксијаоејан, 5000–4200 БП) 100,0% (=12/12)
- Дадијанзи (Култура на Долна Ксијаџиадија, 4200–3600 БП) 60,0% (=3/5)
N-CTS4309: две лица идентификувани со оваа подгрупа во Ирак. Многу ретко.
N1a (F1206/M2013/S11466)
Се проценува дека кладата N1a2-F1008/L666 и N1a1-M46/Page70/Tat го делат најновиот заеднички предок во N1a-F1206/M2013/S11466 приближно 15.900 [95% CI 13.900, сегашноста [90]1 години[1] или 17.621 [95% CI 14.952 <-> 20.282] п.н.е.[3]
N1a1 (M46/Page70/Tat, L395/M2080)
Сите M46 во базата на податоци на Yfull се M178, што се четвртина помлади од одвојувањето од F1139.[13]
Мутациите што ја дефинираат подкладата N-M46 се M46/Tat и P105. Ова е најчестата подклада на N. Веројатно настанала во североисточно азиско население, бидејќи најстарите антички примероци се во согласност со овој генетски профил.[14][15] Н доживеа сериски тесни грла во Сибир и секундарни експанзии во источна Европа. Хаплогрупата N-M46 е стара приближно 14.000 години.
Во Сибир, хаплогрупата N-M46 достигнува максимална честота од приближно 90% кај Јакутите, турски народ кој живее главно во Република Јакутија (Јакутија). Сепак, N-M46 е присутен со многу помала честота кај многу соседи на Јакутите, како што се Евенкс и Евенс.[16] Исто така, откриено е во 5,9% (3/51) од примерокот на Хмонг Да од Лаос, 2,4% (2/85) од примерокот од Сеул, Јужна Кореја и во 1,4% (1/70) од примерок од Токушима, Јапонија.
Хаплогрупата N-M46 има мала разновидност меѓу Јакутите што укажува на тесно грло на населението или ефект на основач. Ова беше потврдено со студија на античка ДНК која го проследи потеклото на машките лоза Јакути до мала група јавачи од областа Цис-Бајкал.
N-Tat е забележан со многу различна честота во примероците од Шведска. Карлсон и сор. (2006) најде N-Tat во 44,7% (17/38) од примерокот на номади Саами од Јокмок, 19,5% (8/41) од примерокот од Вестерботен, 14,5% (8/55) од примерокот од Упсала, 10,0% (4/40) од примерокот од Готланд, 9,5% (4/42) од примерокот од Вермланд, 7,3% (3/41) од примерокот од Остерготланд/Јонкопинг, 2,4% (1/41) од примерок од Блекинге/Кристијанштад и 2,2% (1/45) од примерокот од Скараборг.
Лапалаинен и сор. (2008) најде N-Tat во 14,4% (23/160) од примерок од Шведска.
Лапалаинен и сор. (2009) најде N-Tat во 15,4% (4/26) од примерокот од Седерманланд, 12,5% (3/24) од примерокот од Västmanland, 12,1% (4/33) од примерокот од Упсала, 7,8% (4/51) од примерокот од Гетеборг, 7,0% (3/43) од примерокот од Норботен, 6,8% (5/73) од примерокот од Скане, 6,6% (15/228) од примерокот од Стокхолм, 6,3 % (3/48) од примерокот од Сиднорланд, 6,3% (2/32) од примерокот од Вестерботен, 6,3% (2/32) од примерокот од Оребро, 5,9% (3/51) од примерокот од Вермланд /Даларна, 5,4% (2/37) од примерокот од Остра Готаланд и 5,1% (2/39) од примерокот од југоисточна Шведска (Калмар, Готланд, Кроноберг и Блекинге). Тие не најдоа никаков пример на N-Tat во нивните примероци од Јонкопинг (0/28), Малме (0/29), Халанд (0/34) или Вестра Геталанд (0/75).
N1a1a (M178)
Подкладата N-M178 е дефинирана со присуство на маркери M178 и P298. N-M178* има повисока просечна честота во Северна Европа отколку во Сибир, достигнувајќи честоти од приближно 60% кај Финците и приближно 40% кај Летонците, Литванците и 35% кај Естонците (Derenko 2007 и Lappalainen 2008).
Мирослава Деренко и нејзините колеги забележаа дека има две подгрупа во оваа хаплогрупа, и двете присутни во Сибир и Северна Европа, со различна историја. Оној што тие го нарекоа N3a1 прво се прошири во јужен Сибир и се прошири во Северна Европа. Во меѓувреме, помладиот подкластер, кој тие го нарекоа N3a2, потекнува од јужен Сибир (веројатно во регионот Бајкал).
Неолитските примероци од областа Бајкал дадоа многу примероци од yDNA N, а еден примерок од Фофоново, Бурјатија, 5000-4000 п.н.е. е меѓу првите Тат примероци во античкиот запис.
Најраните примероци на N1a1a-L708 беа пронајдени во Транс-Бајкал (brn008, N1a1a1*-L708; brn003, N1a1a1a1*-M2126) помеѓу 8.000 и 6.000 п.н.е. Низводните примероци беа пронајдени во Јакутија (N4b2, N1a1a1a1a*-Z1979) и Краснојарскиот крај (kra001, N1a1a1a1a*-L392), помеѓу 5.000 и 4.000 п.н.е.[17][18]
N1a2 (F1008/L666)
N1a2a-M128 и N1a2b-B523/P43 се проценува дека споделуваат најнов заеднички предок во N1a2-F1008/L666 приближно 8.600 [95% CI 7.500 <-> 9.800] п.н.е.,[1][19] 9.200 п.н.е.[19] или 9.314 [95% CI 7.419 <-> 11.264] п.н.е.[3]
Најмалку три од шест тестирани машки примероци од слојот од раниот неолит (ловец-собирач на керамика од пред приближно 7200–6200 години) на археолошкиот локалитет Шаманка во близина на јужниот крај на Бајкалското Езеро, се откриени дека припаѓаат на N1a2-L666.
N1a2a-M128
Оваа подклада е дефинирана со присуството на маркерот M128. N-M128 првпат беше идентификуван во примерок од Јапонија (1/23 = 4,3%) и во примерок од Средна Азија и Сибир (1/184 = 0,5%) во прелиминарното истражување на варијации на Y-DNA низ светот.[20] Последователно, беше пронајден со мала честота кај некои примероци од народот Манчу, народот Сибе, Евенки, Корејци, Хан Кинези, Хуи, Тибетанци, Виетнамци, Бујеи, Казахстанци, Узбеци, Ујгури, Салари, Ту, Монголи, Бузава племето на Калмиците,[21] Какас и Комис.[22]
Откриено е дека голем број од Хан Кинез, Оолден Монгол, Кианг и Тибетанец припаѓаат на сестринска гранка (или гранки) на N-M128 под парагрупата N-F1154*.[23]
Неолитскиот примерок brn002 (~ 5.940 БП) во Транс-Бајкал беше откриен дека е ран огранок возводно од N-M128.[24]
Како резултат на генетско тестирање на кланот Јелу, кралско семејство со потекло од династијата Лиао и Китан, беше откриено дека припаѓа на N-F1998, низводно од N-M128.
N1a2b (P43)
Хаплогрупата N-P43 е дефинирана со присуството на маркерот P43. Се проценува дека е старо приближно 4.000 до 5.500 години (TMRCA 4.510 години,[19] TMRCA 4.700 [95% CI 3.800 <-> 5.600] п.н.е.,[1] или 4.727 [95% CI <3] 6,82 п.н.е.[3]). Многу често е пронајден кај северносамоједните народи, говорителите на об-угрските јазици и северните хакасијци, а исто така е забележан со мала до умерена честота меѓу говорителите на некои други уралски јазици, турски народи, монголски народи, тунгуски народи, и сибирскиот народ Јупик.
Највисоките честоти на N-P43 се забележани кај северозападните сибирски популации: 92% (35/38)[22] во примерок од Нганасан, 78% (7/9)[25][26] во примерок од Енети, 78% (21/27)[27] во примерок од Кантс, 75% (44/59)[22] во примерок од Тундра Ненец, 69% (29/42)[3] во друг примерок од Ненец, 60% (15/25)[28] во примерок од Манси, 57% (64/112) во друг примерок на Кантс, 54% (27/50)[3] во друг примерок од Нганасан, 45% (40/89)[22] во примерок од Форест Ненетс, 38% (18/47)[29] во трет примерок од Ханти и 25% (7/28)[28] во четврти примерок од Ханти. Во Европа, типовите N-P43 имаат најголема честота од 20% кај Волга-Уралското население. Екстремната западна граница на ширењето на N-P43 е Финска, каде што оваа хаплогрупа се јавува само на маргинална честота - 0,4%. Сепак, N-P43 е доста чест кај Вепсите (17,9%), мала финска популација која живее во непосредна близина на Финците, Карелијците и Естонците.[22]
Хаплогрупата N-P43, исто така, е забележана со многу висока честота (26/29 = 89,7% од примерокот од населбата Топанов и 19/22 = 86,4% од примерокот од населбата Мали Спирин) во примероците на Качинс, а Турскојазична етничка група или територијална подгрупа на народот Какас, од округот Ширински во северна Хакасија.[30] Се чини дека постои клин низ Сагаите (друга етничка група која зборува турски јазик, која сега се смета за конституент на народот Хака), со 46,2% (55/119) од Сагаи земени од Уст'–Ес', Есино, Уст'–Чул' и Кизлас населбите од округот Аскиски во централна Какасија кои припаѓаат на хаплогрупата N-P43 наспроти само 13,6% (11/81) од Сагаи земени од примероците од населбите Матур, Анчул', Бол'шаја Сеја и Бутрахти во Таштипски Областа на јужна Какасија припаѓа на оваа хаплогрупа.[30] Сепак, примероците на други истражувачи на луѓе од Кака покажаа само умерени честоти на N-P43 или потенцијални N-P43. Деренко и сор. (2006) испита примерок од Хакаси (n=53) собран во населбите во областите Аскиз, Ширинск, Бејск и Орџоникидзевск во Република Хакас и откри дека 15 од нив (28,3%) припаѓаат на N-LLY22g(xTat). Роотси и сор. (2007) испитуваше примерок од Кака (n =181) и откри дека 31 од нив (17,1%) припаѓаат на N-P43;[22] Ilumäe и др. 2016 повторно тестираше 174 од поединците во овој примерок и откри дека 27 од нив (15,5%) припаѓаат на подкладата N-B478 (азиска/северна самоједска) на N-P43 и 2 од нив (1,1%) припаѓале на N-L1419 (европски/Волга-фински и чувашки) подкладата на N-P43 за вкупно 29 (16,7%) N-P43.[3]
Хаплогрупата N-P43 формира две карактеристични подгрупа на STR хаплотипови, азиски и европски, а вторите претежно се дистрибуирани меѓу народите што зборуваат фино-угрички и сродните популации.
N1a2b1-B478
TMRCA на N-B478 се проценува дека е 3.007 [95% CI 2.171 <-> 3.970] п.н.е.[3] Таа е една од најраспространетите хаплогрупи на Y-DNA меѓу домородните популации на северозападен Сибир: 69,0% (29/42) Ненец, 50,0% (25/50) Нганасан, 22,2% (12/54) Долган од Тајмир, 7,0% (3/43) Селкуп, 1,6% (1/63) Об-Угријан. Исто така е доста распространета меѓу популациите на Централен Сибир, Јужен Сибир и Монголија: 17,9% (17/95) Тувани, 15,5% (27/174) Кака, 13,0% (6/46) Тожу Туванци,[31] 8,7% (2/23) Шор, 8,3% (2/24) Парен, 8,2% (5/61) Алтајец, 5,3% (3/57) Евенк, 5,0% (19/381) Монгол, 4,9% (3/61) Сарт-Калмак (делумни потомци на Монголите од Оират во Киргистан),[31] 4,2% (9/216) Јакут, 2,1% (1/47) Торгут (Монголија),[31] 1,4% (1/69) Дербет (Калмикија),[31] 0,9% (1/111) Бурјат. Географски оддалечен член е пронајден во примерок од Чуваш (1/114 = 0,88%).[3]
Карафет и сор. (2018) забележан N-P63, кој се чини дека е приближно филогенетски еквивалентен на N-B478, во 91,2% (31/34) Нганасан, 63,8% (30/47) Тундра Ненец, 42,7% (35/82) Шумски Ненети, 14,0% (8/57) Долган, 7,0% (9/129) Селкуп, 3,3% (3/91) Евенк, 2,7% (2/75) Монгол, 2,6% (2/78) Коми, 2,5% (2/ 80) Бурјат и 2,0% (2/98) Алтај Кижи. Оваа хаплогрупа не беше забележана во примероците на Јукагир (0/10), Корјак (0/11), Телеут (0/40), Кет (0/44), Јакут (0/62) или Канти (0/165) популации.
N1b (F2930)
Хаплогрупата N1b е претежно пронајдена кај народот Ји, етничка група која зборува тибето-бурма во југозападна Кина, која потекнува од древните племиња Чианг во северозападна Кина. Сепак, исто така е пронајден кај луѓе ширум Кина (каде што сочинуваат приближно 3,62% од машката популација во земјата и главно се дистрибуирани во Шандонг, Џиангсу, Жеџијанг, Анхуи итн.[32]) и кај некои поединци од Шпанија, Еквадор, Полска, Белорусија, Русија, Ирак, Индија, Казахстан, Кореја, Јапонија, Бутан, Виетнам, Камбоџа, Лаос, Тајланд, Малезија и Сингапур.[1][9]
N2 (Y6503)
N2 (Y6503/FGC28528; B482/FGC28394/Y6584) - примарна гранка на хаплогрупата N-M231, сега е претставена главно со подклада, N-FGC28435, која се проширила веројатно некое време во првата половина на вториот милениум[33] и тоа е пронајдено кај поединци од Србија, Хрватска, Босна и Херцеговина, Црна Гора и Турција (Истанбул).[33][34]
N-Y7310 (или N-F14667) го сублимира N-FGC28435 и, исто така, веројатно потекнува од заеднички предок кој живеел некое време во првата половина на минатиот милениум. Сепак, членовите на N-Y7310 (xFGC28435) покажуваат поголем географски опсег, вклучувајќи поединец од Ростовската област на Русија и романски унгарски поединец со потекло од Сучеава, Буковина.[1]
Други ограноци на N-P189 вклучуваат членови од Турција,[1] Русија (Московска област[1]), Франција (Charente-Maritime[1]) и Англија (Девон[1]).[9] Најновиот заеднички предок на сите гореспоменатите постоечки лози на N-P189 се проценува дека е 4.900 (95% CI 5.700 <-> 4.100) п.н.е.[1] Археолошки примерок припишан на културата Ботаи во северен Казахстан и датиран во втората половина на четвртиот милениум п.н.е. припаѓа на N-P189*, што е базален на денешните европски членови на N-P189.[1][35]
Лозите кои припаѓаат на N-Y6503(xP189) и се само далечно поврзани (со време на најновиот заеднички предок што се проценува дека е поголемо од 10.000 п.н.е.[1]) со гореспоменатите членови на N-P189 се пронајдени во поединец од денешна Алтајска република[1] и веројатно, исто така, во археолошки примерок припишан на културата Мезочат од железното време на она што е сега Унгарија (приближно 2.900 п.н.е.)[36] и во археолошки примерок припишан на Китои култура на фуражи кои користат керамика од областа околу Бајкалското Езеро (приближно 6700 години пред сега).[35]
Антички народи
Примерок ископан на локацијата Хоутамуга во населбата Југе во градот Хонггангзи, Даан, Џилин, Кина, кој датира од пред 7430-7320 години (II фаза од раниот неолит) е откриено дека припаѓа на Y-DNA хаплогрупата N и mtDNA хаплогрупа B4c1a2. Овој примерок е автосомно идентичен со популациите на неолитскиот басен на реката Амур, од кои жителите на Нивк се најблискиот модерен претставник. Бидејќи трудот го открил ова потекло во терминалниот плеистоцен USR1 примерок во Алјаска, затоа се претпоставува дека постоел проток на гени од Амур во Америка на популација која припаѓа на хипотетичкото јазично семејство Чукотко-Камчаткан-Нивк.
N, исто така, е пронајден во многу примероци од неолитски човечки останки ексхумирани од цивилизацијата Лиао во североисточна Кина и во областа околу Бајкал во јужен Сибир.[37] Се сугерира дека yDNA N, стигна до јужен Сибир од 12 до 14 кв. Од таму стигна до јужна Европа 8-10kya.
Филогенијата
Филогенетско дрво
Во следното стебло, номенклатурата на три извори е одделена со коси црти:
- NO-M214
- N-M231/Page91, M232/M2188
- N1-Z4762/CTS11499/L735/M2291
- N1a-L729
- N-Z1956
- N-Y149447
- N1a1-M46/Page70/Tat
- N1a1a-M178
- N1a1a1-F1419
- N1a1a1a-L708
- N1a1a1a1-P298/M2126
- N1a1a1a1a-L392
- N1a1a1a1a1-CTS10760
- N1a1a1a1a1a-CTS2929/VL29
- N1a1a1a1a1a1-Z4908
- N-Y46443
- N-Y46443*
- N-BY33095
- N1a1a1a1a1a1a-L550/S431
- N-L550*
- N-Y9454
- N-Y20911
- N-Y7795
- N-Y7795*
- N-Y29766
- N-Y20918
- N-Y28771
- N-Y61225
- N-Y30126
- N-Y30126*
- N-Y29764
- N-Y29764*
- N-Y30123
- N-S9378
- N-S9378*
- N-S18447
- N-Y36282
- N-Y36282*
- N-BY21957
- N-FGC14542
- N-FGC14542*
- N-BY21938
- N-Y17113
- N-Y4341
- N-BY21874
- N-Y4338
- N-Y4338*
- N-Y4339
- N-Y12104
- N-Y12104*
- N-Y12103
- N-Y19111
- N-Y57577
- N-Y22774
- N-Y5611
- N-Y5611*
- N-Y21546
- N-F1983
- N-Y10932
- N-Y85136
- N-Y10931
- N-Y12104
- N1a1a1a1a1a1a1-L1025/B215
- N-L1025*
- N-BY30389
- N-Y13982
- N-Y13982*
- N-Y31236
- N-A11940
- N-Y140872
- N-Y143451
- N-Y5580
- N-Y93996
- N-BY158
- N-Z16975
- N-VL69
- N-Z16976
- N-Z16976*
- N-Y13475
- N-Y21578
- N-Y6129
- N-Y19113
- N-Y134492
- N-L551
- N-L551*
- N-Y15251
- N-Y46313
- N-Y86578
- N-Y14152
- N-BY21911
- N-Y13979
- N-Y4706
- N-Y4706*
- N-A705
- N-BY21893
- N-Y139030
- N-Y183040
- N-Y4707
- N-Z16981
- N-A2358
- N-CTS8173
- N-BY32524
- N-FGC39882
- N-A11470
- N-Y15922
- N-Y6075
- N-Y11882
- N-Y11882*
- N-ZS11617
- N-Y24601
- N-Y94659
- N-Y32725
- N-FT96305
- N-Y33333
- N-Y46443
- N1a1a1a1a1a2-CTS9976
- N-L1022
- N-Y19098
- N-Y19098*
- N-A12258
- N-BY117178
- N-Y5004
- N-Y7300
- N-A17632
- N-Y128024
- N-A16017
- N-A16017*
- N-A16526
- N-FT216144
- N-Y95406
- N-Y7297
- N-Y15813
- N-Y15813*
- N-Y49008
- N-Y46886
- N-Y15812
- N-A14187
- N-Y24617
- N-Y23576
- N-Y23576*
- N-Y15615
- N-BY100801
- N-A17838
- N-Y23179
- N-A17632
- N-Y5005
- N-Y22106
- N-Y47789
- N-Y79341
- N-Y10756
- N-A13656
- N-PH2196
- N-CTS11122
- N-A17277
- N-PH547
- N-Y5003
- N-Y5003*
- N-BY22001
- N-BY6007
- N-Y132182
- N-Y20917
- N-Y24502
- N-Y18420
- N-Z35267
- N-Y6599
- N-Y24000
- N-Y24001
- N-A17082
- N-Y16503
- N-Y22106
- N-Y7300
- N-Y19098
- N-BY6010
- N-CTS3451
- N-CTS3451*
- N-Y3667
- N-BY33087
- N-BY33087*
- N-BY33088
- N-CTS657
- N-CTS657*
- N-BY70437
- N-BY6024
- N-Y45925
- N-Y26750
- N-Y26750*
- N-A16653
- N-PH3568
- N-BY33087
- N-L1022
- N1a1a1a1a1a1-Z4908
- N1a1a1a1a1b-PH1266/Y28526/F4134
- N-Y46746
- N-Y32732
- N-Y32732*
- N-Y192174
- N-Y32731
- N1a1a1a1a1c-B479
- N1a1a1a1a1a-CTS2929/VL29
- N1a1a1a1a2-Z1936,CTS10082
- N1a1a1a1a2a1c-PH3340/Y13850
- N1a1a1a1a2a1c1-L1034
- N-Y28538
- N-Y28538*
- N-L1032
- N-L1442
- N-FT12605
- N-Y23732
- N-Y24222
- N-Y24222*
- N-Y62987
- N-Y28538
- N1a1a1a1a2a1c2-Y24361
- N-Y24361*
- N-Y24360
- N1a1a1a1a2a1c1-L1034
- N1a1a1a1a2a-Z1934
- N-Y159520
- N-Y159520*
- N-BY184755
- N-Y18421
- N-Y18421*
- N-Y19110
- N-Y180247
- N-Y180247*
- N-FT52835
- N-Y19108
- N-Y19108*
- N-Y19453
- N-Y19453*
- N-Y19451
- N-Y180247
- N-Z1928/CTS2733
- N-YP6091
- N-YP6091*
- N-YP6094
- N-Y129131
- N-Y129131*
- N-A25107
- N-YP6092
- N-YP6092*
- N-B195
- N-B195*
- N-Y62142
- N-Y129131
- N1a1a1a1a2a-Z1925
- N-Z1925*
- N-Y29767
- N-Y62904
- N1a1a1a1a2a2a1a1-Z1926
- N-Y20920
- N-Y20920*
- N-Y21699
- N-Z1927
- N-Y22108
- N-Z1933
- N-CTS4329
- N-Y22091
- N-Y22091*
- N-Y47623
- N-Y47623*
- N-Y106419
- N-Y31247
- N-Y31247*
- N-Y31249
- N-Y90283
- N-Y31244
- N-YP6269
- N-YP6269*
- N-Y151660
- N-Y151660*
- N-BY149208
- N-CTS8565
- N-BY18114
- N-BY30394
- N-Y30513
- N-Y30513*
- N-Y29759
- N-Y29759*
- N-Y29758
- N-Z4998
- N-Z4998*
- N-FGC65190
- N-Y18414
- N-Y20910
- N-BY194138
- N-Y28547
- N-BY22141
- N-FT5834
- N-BY190112
- N-Y19097
- N-CTS3223
- N-Y22091
- N-VL62
- N-VL62*
- N-VL60
- N-VL60*
- N-Y63781
- N-VL72
- N-VL72*
- N-BY30470
- N-Y20915
- N-Z1939
- N-Y132561
- N-Y132565
- N-Y152012
- N-Y152012*
- N-Y152010
- N-Z2445
- N-Z35031
- N-BY28931
- N-Y125841
- N-Y11631
- N-CTS7189
- N-BY142665
- N-Y37149
- N-Y23568
- N-Y80226
- N-FT20730
- N-BY62666
- N-Y24218
- N-Y24218*
- N-Y24217
- N-BY22038
- N-Y13974
- N-Y135981
- N-Y132561
- N-Z4747
- N-Y17790
- N-Z1941
- N-BY22090
- N-Y21575
- N-Z1940
- N-CTS4329
- N-Y20920
- N-YP6091
- N-Y159520
- N1a1a1a1a2a1c-PH3340/Y13850
- N1a1a1a1a3-B197/Y16323
- N1a1a1a1a3a-F4205
- N-F4205*
- N-Y16221
- N-Y16221*
- N-Y16220
- N-Y16220*
- N-F22331
- N-Y63966
- N-Y226011
- N-Z35331
- N-B199
- N-FT411781
- N-Y16312/F2288
- N-Y16312*
- N-Y16320
- N-Z35331
- N1a1a1a1a3b-B202
- N1a1a1a1a3a-F4205
- N1a1a1a1a1-CTS10760
- N1a1a1a1b-M2019/M2118
- N-M2058
- N-M2016
- N-M1993
- N-FT411801
- N1a1a1a1b1-M2038/M1995
- N-M1932
- N-M1991
- N-M1988
- N-Y25011
- N-M1993
- N1a1a1a1b2-A9408
- N-Y70200
- N-Y60223
- N-PH1612
- N-A9407/A9411
- N-A9416
- N-PH1896
- N-A9407/A9411
- N-Y70200
- N-M2016
- N-M2058
- N1a1a1a1a-L392
- N1a1a1a2-B211 (Y9022)
- N1a1a1a2a-B181
- N-Y182299
- N-Y23788
- N1a1a1a2b-Y23181
- N-Y23183
- N-Y143277
- N1a1a1a2a-B181
- N1a1a1a1-P298/M2126
- N-Y24317
- N-Y24317*(xB187)
- N1a1a1b-B187
- N1a1a1a-L708
- N1a1a2-Y23747
- N1a1a2a-Y23749
- N1a1a2b-Y125664
- N-Y125664*
- N-MF16376
- N1a1a2b1-MF38607
- N1a1a2b2-F22150
- N-FT281705
- N-MF200125
- N-MF43738
- N-MF82671
- N-MF124520
- N-MF58064/F19130
- N-MF82671
- N-MF15288/MF15344
- N-MF55680
- N-MF37603
- N-F21121
- N-MF48201
- N-FT281705
- N1a1a1-F1419
- N1a1a-M178
- N-F1360
- N-F1360*
- N1a2-F1008/L666
- N-F1101
- N-F1101*
- N-F1154
- N-F1154*
- N-Y23741
- N-Y23741*
- N1a2a-M128
- N-M128*
- N-Y23738
- N-CTS1350
- N-CTS1350*
- N-F1998
- N-F1998*
- N-MF1633
- N-MF41795
- N-Y71703
- N-Y71703*
- N-Y173843
- N-Y173843*
- N-Y174089
- N1a2b-B523(P43)
- N1a2b-Y126204, VL67/Z35079, BY29083
- N-VL67/Z35079
- N1a2b1-B478 (P63)
- N1a2b1a-B168
- N1a2b1b-B169
- N1a2b1b1-B170
- N1a2b1b2-B175
- N1a2b3-B525
- N-B525*
- N-BY173686
- N-VL83
- N-Y136502
- N-Y37153
- N-Y37153*
- N-BY29131
- N-BY29131*
- N-BY30476
- N1a2b1-B478 (P63)
- N1a2b2-FGC10872/Y3195
- N1a2b2a-FGC10847/Y3185 (L1419)
- N1a2b2b-Y23786
- N1a2b2b*
- N1a2b2b1-B528/Y24384
- N-VL67/Z35079
- N1a2b-Y126204, VL67/Z35079, BY29083
- N-F1101
- N-Z1956
- N1b-F2905
- N1b1-CTS582
- N1b1a-Y6374/Z8029
- N1b1a1-CTS7324
- N1b1a2-L727
- N1b1a2*-L727
- N1b1a2a-L732
- N1b1a2a1-F839
- N-F18335
- N-F16160
- N-Y137601
- N-F18335
- N1b1a2a2-Y15972
- N-F21018/MF105942
- N-Y62279
- N-MF155949
- N-L733
- N-Y154812
- N-Y16106
- N-Y232856
- N-Y15965
- N-Y232856
- N-Y16106
- N-Y154812
- N-F21018/MF105942
- N1b1a2a1-F839
- N1b1b-Y23789/CTS4309
- N1b1b*-Y23789
- N1b1b1-Y60861
- N1b1a-Y6374/Z8029
- N1b2-M1819/N-M1897/CTS12473/F1173
- N-M1897*
- N-M1845
- N-M1845*
- N-M1928
- N-M1928*
- N-Y125475
- N-Y63516
- N-Y193396
- N-CTS4714
- N-CTS4714*
- N-F2407
- N-Y24191
- N-Y24191*
- N-Y24193
- N-Y24193*
- N-Y24190
- N-M1877
- N-M1877*
- N-F1486
- N-F1486*
- N-M1812
- N-F1260
- N-F1260*
- N-Y62067
- N-Y62067*
- N-Y62132
- N-M1823
- N-M1823*
- N-M1811
- N-M1811*
- N-Y24355
- N-Y24355*
- N-Y64234
- N-F1260
- N-Y24191
- N1b1-CTS582
- N1a-L729
- N2-Y6503
- N2-Y6503*
- N2a-P189.2
- N2a1-Y6516
- N2a1-Y6516*
- N2a1a-Y7310
- N2a1a-Y7310*
- N2a1a1-Y7313
- N2a1a1-Y7313*
- N2a1a1a-BY35494
- N2a1a1b-FGC28435
- N2a1a1b-FGC28435*
- N2a1a1b1-FGC28483
- N2a2-Y111068
- N2a2a-FT352925 [38]
- N2a2b-Y101945
- N2a1-Y6516
- N1-Z4762/CTS11499/L735/M2291
- N-M231/Page91, M232/M2188
Историја на филогенетската номенклатура
Пред 2002 година, во академската литература имаше најмалку седум системи за именување на филогенетското дрво на Y-хромозомот. Ова доведе до значителна конфузија. Во 2002 година, главните истражувачки групи се собраа и го формираа Y-хромозомот конзорциум (YCC). Тие објавија заеднички труд кој создаде единствено ново дрво кое сите се согласија да го користат. Подоцна, група граѓански научници со интерес за популациската генетика и генетската генеалогија формираа работна група за да се создаде аматерско дрво со цел да биде пред сè навремено. Табелата подолу ги обединува сите овие дела на точката на знаменитото дрво на МКЦ од 2002 година. Ова му овозможува на истражувачот што прегледува постара објавена литература брзо да се движи помеѓу номенклатурите.
| МКЦ 2002/2008 (стенограф) | (α) | (β) | (γ) | (δ) | (ε) | (ζ) | (η) | МКЦ 2002 година | МКЦ 2005 година | МКЦ 2008 година | МКЦ 2010r | ISOGG 2006 година | ISOGG 2007 година | ISOGG 2008 година | ISOGG 2009 година | ISOGG 2010 година | ISOGG 2011 година | ISOGG 2012 година |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N-LLY22g | 12 | VIII | 1U | 25 | ЕУ16 | Н5 | Ф | N* | Н | N1 | N1 | - | - | - | - | - | - | - |
| N-M128 | 12 | VIII | 1U | 25 | ЕУ16 | Н5 | Ф | N1 | N1 | N1a | N1a | - | - | - | - | - | - | - |
| N-P63 | 12 | VIII | 1U | 25 | ЕУ16 | Н5 | Ф | N2 | N2a | N1b1 | N1b1 | - | - | - | - | - | - | - |
| Н-ТАТ | 12 | VIII | 1I | 26 | ЕУ13 | Н5 | Ф | N3* | N3 | N1c | N1c | - | - | - | - | - | - | - |
| Н-М178 | 16 | VIII | 1I | 26 | ЕУ14 | Н5 | Ф | N3a* | М178 | N1c1 | N1c1 | - | - | - | - | - | - | - |
| N-P21 | 16 | VIII | 1I | 26 | ЕУ14 | Н5 | Ф | N3a1 | N3a1 | N1c1a | N1c1a | - | - | - | - | - | - | - |
Извори Следниве истражувачки тимови по нивните публикации беа претставени во креирањето на Дрвото на МКЦ.
Несигурни мутации (SNP и UEPs)
Бришењето b2/b3 во AZFc регионот на Y-хромозомот се чини дека се случило независно во најмалку четири различни прилики. Затоа, ова бришење не треба да се земе како уникатен настан полиморфизам кој ја дефинира оваа гранка на дрвото на Y-хромозомот ISOGG 2012.
Наводи
- ↑ 1,00 1,01 1,02 1,03 1,04 1,05 1,06 1,07 1,08 1,09 1,10 1,11 1,12 1,13 1,14 1,15 1,16 1,17 YFull Haplogroup YTree v6.05.11 at 25 September 2018.
- ↑ Poznik GD, Xue Y, Mendez FL, Willems TF, Massaia A, Wilson Sayres MA, и др. (June 2016). „Punctuated bursts in human male demography inferred from 1,244 worldwide Y-chromosome sequences“. Nature Genetics. 48 (6): 593–599. doi:10.1038/ng.3559. PMC 4884158. PMID 27111036.
- ↑ 3,00 3,01 3,02 3,03 3,04 3,05 3,06 3,07 3,08 3,09 Ilumäe и др. 2016
- ↑ 4,0 4,1 4,2 ISOGG, 2016, Y-DNA Haplogroup N and its Subclades – 2016 22 August 2016).
- ↑ Rootsi и др. 2006
- ↑ „Genetic evidence of an East Asian origin and paleolithic northward migration of Y-chromosome haplogroup N“. PLOS ONE. 8 (6): e66102. 2013. Bibcode:2013PLoSO...866102S. doi:10.1371/journal.pone.0066102. PMC 3688714. PMID 23840409.
- ↑ „A recent bottleneck of Y chromosome diversity coincides with a global change in culture“. Genome Research. 25 (4): 459–466. April 2015. doi:10.1101/gr.186684.114. PMC 4381518. PMID 25770088.CS1-одржување: display-автори (link)
- ↑ „The first horse herders and the impact of early Bronze Age steppe expansions into Asia“. Science. 360 (6396): eaar7711. June 2018. doi:10.1126/science.aar7711. PMC 6748862. PMID 29743352.CS1-одржување: display-автори (link)
- ↑ 9,0 9,1 9,2 Tree of Y-DNA haplogroup N-M231 at Family Tree DNA Discover
- ↑ „Haplotree.info - ancientdna.info. Map based on All Ancient DNA v. 2.07.26“.
- ↑ Wen и др. 2004.
- ↑ „Y Chromosome analysis of prehistoric human populations in the West Liao River Valley, Northeast China“. BMC Evolutionary Biology. 13: 216. September 2013. doi:10.1186/1471-2148-13-216. PMC 3850526. PMID 24079706.CS1-одржување: display-автори (link)
- ↑ „N-Z1956 YTree“.
- ↑ „Human auditory ossicles as an alternative optimal source of ancient DNA“. Genome Research. 30 (3): 427–436. March 2020. doi:10.1101/gr.260141.119. PMC 7111520. PMID 32098773.CS1-одржување: display-автори (link)
- ↑ „Transcriptional profiling of lung cell populations in idiopathic pulmonary arterial hypertension“. Pulmonary Circulation. 10 (1). 2020. doi:10.1101/2020.10.12.336628. PMC 7052475. PMID 32166015.CS1-одржување: display-автори (link)
- ↑ „Investigating the prehistory of Tungusic peoples of Siberia and the Amur-Ussuri region with complete mtDNA genome sequences and Y-chromosomal markers“. PLOS ONE. 8 (12): e83570. 2013. Bibcode:2013PLoSO...883570D. doi:10.1371/journal.pone.0083570. PMC 3861515. PMID 24349531.CS1-одржување: display-автори (link)
- ↑ „Human population dynamics and Yersinia pestis in ancient northeast Asia“. Science Advances. 7 (2): eabc4587. January 2021. Bibcode:2021SciA....7.4587K. doi:10.1126/sciadv.abc4587. PMC 7787494 Проверете ја вредноста
|pmc=(help). PMID 33523963 Проверете ја вредноста|pmid=(help).CS1-одржување: display-автори (link) - ↑ „N-L708 YTree“. www.yfull.com. Посетено на 2022-11-28.
- ↑ 19,0 19,1 19,2 Phylogenetic tree of Haplogroup N at 23mofang
- ↑ Underhill и др. 2000.
- ↑ „Y-chromosome diversity in the Kalmyks at the ethnical and tribal levels“. Journal of Human Genetics. 58 (12): 804–811. December 2013. doi:10.1038/jhg.2013.108. PMID 24132124.
- ↑ 22,0 22,1 22,2 22,3 22,4 22,5 Rootsi и др. 2006.
- ↑ Kang Hu и др. 2015.
- ↑ „Ancient Y-DNA with reconstructed phylogeny provides insights into the demographic history of paternal haplogroup N1a2-F1360“. Journal of Genetics and Genomics = Yi Chuan Xue Bao. 48 (12): 1130–1133. December 2021. doi:10.1016/j.jgg.2021.07.018. PMID 34425243 Проверете ја вредноста
|pmid=(help).CS1-одржување: display-автори (link) - ↑ Tatiana M. Karafet, Ludmila P. Osipova, Marina A. Gubina, Olga L. Posukh, Stephen L. Zegura, and Michael F. Hammer, "High Levels of Y-Chromosome Differentiation among Native Siberian Populations and the Genetic Signature of a Boreal Hunter-Gatherer Way of Life." Human Biology, December 2002, v. 74, no. 6, pp. 761–789.
- ↑ „Genes reveal traces of common recent demographic history for most of the Uralic-speaking populations“. Genome Biology. 19 (1): 139. September 2018. doi:10.1186/s13059-018-1522-1. PMC 6151024. PMID 30241495.CS1-одржување: display-автори (link)
- ↑ „Y-chromosome distribution within the geo-linguistic landscape of northwestern Russia“. European Journal of Human Genetics. 17 (10): 1260–1273. October 2009. doi:10.1038/ejhg.2009.6. PMC 2986641. PMID 19259129.CS1-одржување: display-автори (link)
- ↑ 28,0 28,1 „Northwest Siberian Khanty and Mansi in the junction of West and East Eurasian gene pools as revealed by uniparental markers“. European Journal of Human Genetics. 16 (10): 1254–1264. October 2008. doi:10.1038/ejhg.2008.101. PMID 18506205.
- ↑ „The western and eastern roots of the Saami--the story of genetic "outliers" told by mitochondrial DNA and Y chromosomes“. American Journal of Human Genetics. 74 (4): 661–682. April 2004. doi:10.1086/383203. PMC 1181943. PMID 15024688.CS1-одржување: display-автори (link)
- ↑ 30,0 30,1 „Transcriptional profiling of lung cell populations in idiopathic pulmonary arterial hypertension“. Pulmonary Circulation. 10 (1): 404–416. 2011. doi:10.1134/S0026893311020117. PMC 7052475. PMID 32166015.CS1-одржување: display-автори (link)
- ↑ 31,0 31,1 31,2 31,3 „Y-chromosomal analysis of clan structure of Kalmyks, the only European Mongol people, and their relationship to Oirat-Mongols of Inner Asia“. European Journal of Human Genetics. 27 (9): 1466–1474. September 2019. doi:10.1038/s41431-019-0399-0. PMC 6777519. PMID 30976109.CS1-одржување: display-автори (link)
- ↑ „N-F2930单倍群详情“.
- ↑ 33,0 33,1 N-P189.2 YFull, 2018, (24 June 2018).
- ↑ Y-DNA Haplogroup N and its Subclades – 2018, ISOGG, 2018, (24 June 2018).
- ↑ 35,0 35,1 Peter de Barros Damgaard, Nina Marchi, Simon Rasmussen, et al. (2018), "137 ancient human genomes from across the Eurasian steppes." Nature, volume 557, pages 369–374 (2018).
- ↑ „Genome flux and stasis in a five millennium transect of European prehistory“. Nature Communications. 5: 5257. October 2014. Bibcode:2014NatCo...5.5257G. doi:10.1038/ncomms6257. PMC 4218962. PMID 25334030.CS1-одржување: display-автори (link)
- ↑ „Y Chromosome analysis of prehistoric human populations in the West Liao River Valley, Northeast China“. BMC Evolutionary Biology. 13 (1): 216. September 2013. doi:10.1186/1471-2148-13-216. PMC 3850526. PMID 24079706.CS1-одржување: display-автори (link)
- ↑ Предупредување за наводот: Ознаката
<ref>со називYFull2не може да се прегледа бидејќи е определена вон тековниот оддел, или пак воопшто не е определена.
Библиографија
- Cai X, Qin Z, Wen B, Xu S, Wang Y, Lu Y, и др. (2011). O'Rourke D (уред.). „Human migration through bottlenecks from Southeast Asia into East Asia during Last Glacial Maximum revealed by Y chromosomes“. PLOS ONE. 6 (8): e24282. Bibcode:2011PLoSO...624282C. doi:10.1371/journal.pone.0024282. PMC 3164178. PMID 21904623.
- Chiaroni J, Underhill PA, Cavalli-Sforza LL (December 2009). „Y chromosome diversity, human expansion, drift, and cultural evolution“. Proceedings of the National Academy of Sciences of the United States of America. 106 (48): 20174–20179. Bibcode:2009PNAS..10620174C. doi:10.1073/pnas.0910803106. PMC 2787129. PMID 19920170.
- Crubézy E, Amory S, Keyser C, Bouakaze C, Bodner M, Gibert M, и др. (January 2010). „Human evolution in Siberia: from frozen bodies to ancient DNA“. BMC Evolutionary Biology. 10: 25. doi:10.1186/1471-2148-10-25. PMC 2829035. PMID 20100333.
- Derenko M, Malyarchuk B, Denisova G, Wozniak M, Grzybowski T, Dambueva I, Zakharov I (2007). „Y-chromosome haplogroup N dispersals from south Siberia to Europe“. Journal of Human Genetics. 52 (9): 763–770. doi:10.1007/s10038-007-0179-5. PMID 17703276.
- Gayden T, Cadenas AM, Regueiro M, Singh NB, Zhivotovsky LA, Underhill PA, и др. (May 2007). „The Himalayas as a directional barrier to gene flow“. American Journal of Human Genetics. 80 (5): 884–894. doi:10.1086/516757. PMC 1852741. PMID 17436243.
- Hammer MF, Karafet TM, Park H, Omoto K, Harihara S, Stoneking M, Horai S (2005). „Dual origins of the Japanese: common ground for hunter-gatherer and farmer Y chromosomes“. Journal of Human Genetics. 51 (1): 47–58. doi:10.1007/s10038-005-0322-0. PMID 16328082.
- Ilumäe AM, Reidla M, Chukhryaeva M, Järve M, Post H, Karmin M, и др. (July 2016). „Human Y Chromosome Haplogroup N: A Non-trivial Time-Resolved Phylogeography that Cuts across Language Families“. American Journal of Human Genetics. 99 (1): 163–173. doi:10.1016/j.ajhg.2016.05.025. PMC 5005449. PMID 27392075.
- Kang Hu; Shi Yan; Kai Liu; Chao Ning; Lan-Hai Wei; Shi-Lin Li; Bing Song; Ge Yu; Feng Chen; Li-Jun Liu; Zhi-Peng Zhao; Chuan-Chao Wang; Ya-Jun Yang; Zhen-Dong Qin; Jing-Ze Tan; Fu-Zhong Xue; Hui Li; Long-Li Kang; Li Jin (2015). „The dichotomy structure of Haplogroup N“. arXiv:1504.06463 [q-bio.PE].
- Karafet T, Xu L, Du R, Wang W, Feng S, Wells RS, и др. (September 2001). „Paternal population history of East Asia: sources, patterns, and microevolutionary processes“. American Journal of Human Genetics. 69 (3): 615–628. doi:10.1086/323299. PMC 1235490. PMID 11481588. In this article, the "Southern Han" sample of Karafet and Hammer's research group is described as originating from Guangdong, and the "Northern Han" sample is described as originating from Shaanxi.
- Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura SL, Hammer MF (May 2008). „New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree“. Genome Research. 18 (5): 830–838. doi:10.1101/gr.7172008. PMC 2336805. PMID 18385274.
- Karafet TM, Hallmark B, Cox MP, Sudoyo H, Downey S, Lansing JS, Hammer MF (August 2010). „Major east-west division underlies Y chromosome stratification across Indonesia“. Molecular Biology and Evolution. 27 (8): 1833–1844. doi:10.1093/molbev/msq063. PMID 20207712.
- Katoh T, Munkhbat B, Tounai K, Mano S, Ando H, Oyungerel G, и др. (February 2005). „Genetic features of Mongolian ethnic groups revealed by Y-chromosomal analysis“. Gene. 346: 63–70. doi:10.1016/j.gene.2004.10.023. PMID 15716011.
- Khar'kov VN, Stepanov VA, Medvedeva OF, Spiridonova MG, Voevoda MI, Tadinova VN, Puzyrev VP (May 2007). „[Gene pool differences between northern and southern Altaians inferred from the data on Y-chromosomal haplogroups]“. Genetika. 43 (5): 675–687. doi:10.1134/S1022795407050110. PMID 17633562. S2CID 566825.
- Kharkov, V. N.; Stepanov, V. A.; Medvedeva, O. F.; Spiridonova, M. G.; Voevoda, M. I.; Tadinova, V. N.; Puzyrev, V. P. (2007). „Gene Pool Differences between Northern and Southern Altaians Inferred from the Data on Y-Chromosomal Haplogroups“. Genetika. 43 (5): 675–687. doi:10.1134/S1022795407050110. PMID 17633562. S2CID 566825.
- Kim W, Yoo TK, Kim SJ, Shin DJ, Tyler-Smith C, Jin HJ, и др. (January 2007). Blagosklonny M (уред.). „Lack of association between Y-chromosomal haplogroups and prostate cancer in the Korean population“. PLOS ONE. 2 (1): e172. Bibcode:2007PLoSO...2..172K. doi:10.1371/journal.pone.0000172. PMC 1766463. PMID 17245448.
- Lappalainen T, Laitinen V, Salmela E, Andersen P, Huoponen K, Savontaus ML, Lahermo P (May 2008). „Migration waves to the Baltic Sea region“. Annals of Human Genetics. 72 (Pt 3): 337–348. doi:10.1111/j.1469-1809.2007.00429.x. PMID 18294359. S2CID 32079904.
- Malyarchuk B, Derenko M, Grzybowski T, Lunkina A, Czarny J, Rychkov S, и др. (December 2004). „Differentiation of mitochondrial DNA and Y chromosomes in Russian populations“. Human Biology. 76 (6): 877–900. doi:10.1353/hub.2005.0021. PMID 15974299. S2CID 17385503.
- Pakendorf B, Morar B, Tarskaia LA, Kayser M, Soodyall H, Rodewald A, Stoneking M (February 2002). „Y-chromosomal evidence for a strong reduction in male population size of Yakuts“. Human Genetics. 110 (2): 198–200. doi:10.1007/s00439-001-0664-4. PMID 11935328. S2CID 30671159.
- Rootsi S, Zhivotovsky LA, Baldovič M, Kayser M, Kutuev IA, Khusainova R, и др. (6 December 2006). „A counter-clockwise northern route of the Y-chromosome haplogroup N from Southeast Asia towards Europe“. European Journal of Human Genetics. 15 (2): 204–211. doi:10.1038/sj.ejhg.5201748. PMID 17149388.
- Sirak K, Fernandes D, Cheronet O, Harney E, Mah M, Mallick S, и др. (March 2020). „Human auditory ossicles as an alternative optimal source of ancient DNA“. Genome Research. 30 (3): 427–436. doi:10.1101/gr.260141.119. PMC 7111520. PMID 32098773.
- Wen B, Xie X, Gao S, Li H, Shi H, Song X, и др. (May 2004). „Analyses of genetic structure of Tibeto-Burman populations reveals sex-biased admixture in southern Tibeto-Burmans“. American Journal of Human Genetics. 74 (5): 856–865. doi:10.1086/386292. PMC 1181980. PMID 15042512.
- Xue Y, Zerjal T, Bao W, Zhu S, Shu Q, Xu J, и др. (April 2006). „Male demography in East Asia: a north-south contrast in human population expansion times“. Genetics. 172 (4): 2431–2439. doi:10.1534/genetics.105.054270. PMC 1456369. PMID 16489223.
Веб-страници
- ISOGG (2012). „Y-DNA Haplogroup Tree 2012“.
- „Macdonald Y Haplogroups of the World“ (PDF). 2004. Архивирано од изворникот (PDF) на 2004-07-28. Посетено на 2009-08-29.
Извори за табели за конверзија
- Capelli, Cristian; Wilson, James F.; Richards, Martin; Stumpf, Michael P.H.; и др. (February 2001). „A Predominantly Indigenous Paternal Heritage for the Austronesian-Speaking Peoples of Insular Southeast Asia and Oceania“. The American Journal of Human Genetics. 68 (2): 432–443. doi:10.1086/318205. PMC 1235276. PMID 11170891.
- Hammer, Michael F.; Karafet, Tatiana M.; Redd, Alan J.; Jarjanazi, Hamdi; и др. (1 July 2001). „Hierarchical Patterns of Global Human Y-Chromosome Diversity“. Molecular Biology and Evolution. 18 (7): 1189–1203. doi:10.1093/oxfordjournals.molbev.a003906. PMID 11420360.
- Jobling, Mark A.; Tyler-Smith, Chris (2000), „New uses for new haplotypes“, Trends in Genetics, 16 (8): 356–62, doi:10.1016/S0168-9525(00)02057-6, PMID 10904265
- Kaladjieva, Luba; Calafell, Francesc; Jobling, Mark A; Angelicheva, Dora; и др. (February 2001). „Patterns of inter- and intra-group genetic diversity in the Vlax Roma as revealed by Y chromosome and mitochondrial DNA lineages“. European Journal of Human Genetics. 9 (2): 97–104. doi:10.1038/sj.ejhg.5200597. PMID 11313742.
- Semino O, Passarino G, Oefner PJ, Lin AA, и др. (2000), „The Genetic Legacy of Paleolithic Homo sapiens sapiens in Extant Europeans: A Y Chromosome Perspective“, Science, 290 (5494): 1155–1159, Bibcode:2000Sci...290.1155S, doi:10.1126/science.290.5494.1155, PMID 11073453
- Su, Bing; Xiao, Junhua; Underhill, Peter; Deka, Ranjan; и др. (December 1999). „Y-Chromosome Evidence for a Northward Migration of Modern Humans into Eastern Asia during the Last Ice Age“. The American Journal of Human Genetics. 65 (6): 1718–1724. doi:10.1086/302680. PMC 1288383. PMID 10577926.
- Underhill PA, Shen P, Lin AA, Jin L, и др. (November 2000). „Y chromosome sequence variation and the history of human populations“. Nature Genetics. 26 (3): 358–361. doi:10.1038/81685. PMID 11062480. S2CID 12893406.
Дополнителна литература
- ISOGG (2006). „Y-DNA Haplogroup Tree 2006“.
- ISOGG (2007). „Y-DNA Haplogroup Tree 2007“.
- ISOGG (2008). „Y-DNA Haplogroup Tree 2008“.
- ISOGG (2009). „Y-DNA Haplogroup Tree 2009“.
- ISOGG (2010). „Y-DNA Haplogroup Tree 2010“.
- ISOGG (2011). „Y-DNA Haplogroup Tree 2011“.
- ISOGG (2018). „Y-DNA Haplogroup N and its Subclades“.
- YFull. „YFull Experimental YTree“.
- ISOGG 2019/2020: https://docs.google.com/spreadsheets/d/1ju7oNjHjMrgMUB1xXmr0EaaL-RctJJ8FreUPM2DzuBY/edit#gid=692817756
Надворешни врски
- Ширење на Haplogroup N, од The Genographic Project, National Geographic
- N Северноевроазиски YDNA проект во FamilyTreeDNA
- N Y-DNA Хаплогрупа проект во FamilyTreeDNA
- N1c1 Y-DNA Хаплогрупа проект во FamilyTreeDNA
- Хаплогрупата N на Y-хромозомот се распрснува од јужен Сибир до Европа
- Проект за ДНК на династијата Рурикид во FamilyTreeDNA
- Проект за ДНК на руското благородништво во FamilyTreeDNA
| ||||||||||||||