Жабы
| Жабы | |
|---|---|
 Обыкновенная жаба | |
| Научная классификация | |
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Класс: Подкласс: Инфракласс: Надотряд: Прыгающие Отряд: Подотряд: Надсемейство: Hyloidea Семейство: Жабы |
|
| Международное научное название | |
| Bufonidae J. E. Gray, 1825 | |
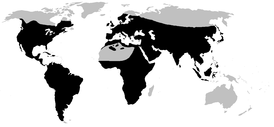
| Ареал | |
 |
|
Жабы[1], или настоящие жабы[1] (лат. Bufonidae), — семейство бесхвостых земноводных, единственное, в котором все представители называются «жабами», хотя некоторые виды можно назвать лягушками из-за похожих внешних черт (например, ателопов). На более высоком таксономическом уровне применение термина «жаба» не ограничивается только данным семейством, а также используется для видов из других семейств (например: жаба-повитуха, седлоносная жаба, лягушковидная жаба, носатая жаба).
Первые представители семейства эволюционно возникли около 80 млн лет назад в Южной Америке и быстро распространились по всему миру. Самые крупные жабы (ага) могут достигать длины 25—50 см и весить более трёх килограммов; наименьшие представители семейства имеют длину не более 2 см. Рёбра отсутствуют. Кожа, как правило, сухая на ощупь, толстая, зачастую покрыта многочисленными гладкими или острыми бугорками-бородавками разного размера; позади глаз хорошо заметны околоушные железы (паротиды), которые у ряда тропических форм отсутствуют. На них содержится яд буфотоксин. Жабы — ночные наземные амфибии, входящие в воду лишь для откладывания яиц, однако существуют и полуводные виды (например, анзонии) и древесные (древесные жабы). Зимуют в земле. Питаются исключительно мелкими беспозвоночными: насекомыми, червями и улитками. Способы размножения разные — от откладывания икры в воду и последующего выхода из неё свободноплавающих личинок, до яйцеживорождения и прямого наземного развития. Большинству видов свойственно внешнее оплодотворение, однако некоторым — внутреннее. На ноябрь 2018 года в семейство включают 52 рода и 609 видов.
Этимология названия
Слово жаба происходит от праславянского *gēbā (ср. бел. жа́ба, укр. жа́ба, болг. жа́ба, сербохорв. жȁба, словен. žába, чеш. žába, слвц. žaba, польск. żaba, в.-луж., н.-луж. žaba, полаб. zobó). На индоевропейском уровне родственно др.-прусск. gabawo «жаба», др.-сакс. quappa, ср.-нидерл. quappe «налим», ср.-нж.-н. quabbe, quobbe «влажная масса», возм., также лат. būfō (ср. научное название жаб)[2].
Область охвата термина
В разговорной речи животных из отряда бесхвостых земноводных называют «лягушками» или «жабами» (возможно провести параллель с латинскими терминами, используя в качестве основы лат. rana для слова «лягушка» и лат. bufo для слова «жаба»). Визуальное отличие одних от других состоит в том, что кожа жаб бородавчатая[3]. При попытке наложить разговорные термины «лягушка» и «жаба» на принятую классификацию отряда бесхвостых оказывается, что практически все семейства бесхвостых включают и тех и других[4].
Эволюция и происхождение
Считается, что первые представители семейства Bufonidae произошли из Южной Америки. Некоторые исследования датируют происхождение группы после распада Гондваны, около 78-98 миллионов лет назад в позднем меловом периоде[5]. Напротив, другие исследования датируют происхождение группы ранним палеоценом[6]. Первые настоящие жабы, вероятно, распространились из Южной Америки в течение эоцена, при этом всё расселение произошло в период от эоцена до олигоцена, что свидетельствует о чрезвычайно быстром расхождении, вероятно, которому способствовали изменяющиеся климатические условия палеогена[6].
Таксономия
Следующая филогения большинства родов семейства основана на исследованиях Portik and Papenfuss, 2015[7], Chan et al., 2016[8], Chandramouli et al. , 2016[9], и Kok et al., 2017[6]:
Описание


Внешность и размеры жаб сильно варьируют. Длина тела колеблется от 20 мм (гвианский арлекин)[10] до 550 мм (жаба-ага)[11]. Масса — от нескольких граммов до 3,65 кг. Туловище обычно широкое, грузное, конечности короткие[12]. Рёбра отсутствуют. Кожа, как правило, сухая на ощупь, толстая, зачастую покрыта многочисленными гладкими или острыми бугорками-бородавками разного размера[13]. Язык узкий, большой. Зубы в большинстве случаев частично или полностью редуцированы. Зрачок горизонтальный, пальцы задних конечностей частично связаны плавательной перепонкой[14]. Поперечные отростки крестцового позвонка не расширены в диски[15]. Настоящие жабы широко распространены и обитают на всех континентах, кроме Австралии и Антарктиды, обитая в самых разных средах, от засушливых районов до тропических лесов. Большинство откладывает яйца парными цепочками, из которых вылупляются головастики, хотя у представителей рода Nectophrynoides прямо из яиц вылупляются миниатюрные жабы[16].
Позади глаз хорошо заметны околоушные железы (паротиды), которые у ряда тропических форм отсутствуют. Эти железы содержат алкалоидный яд — буфотоксин, который жабы выделяют при стрессе. Состав и пропорции яда у разных видов различаются[17]. Яд некоторых жаб (например, колорадской жабы) используется в качестве рекреационного психотропного средства[18].
У самцов семейства присутствует уникальный неотенический орган Биддера — рудиментарный яичник, который развивается на переднем крае личиночных семенников и может сохраняться у многих взрослых амфибий[19]. При определённых условиях орган становится активным яичником, и самец, по сути, становится самкой[20][21][22].
Образ жизни
Питание
Жабы — ночные наземные амфибии, входящие в воду лишь для откладывания яиц, однако существуют и полуводные виды (например, анзонии) и древесные (древесные жабы). Зимуют в земле. Все виды жаб хищные; питаются исключительно мелкими беспозвоночными: насекомыми, червями и улитками. Существуют крупные виды, которые поедают других земноводных, небольших млекопитающих и птиц. Одни ловят свою быстро двигающуюся добычу клейким языком, другие запихивают пищу в рот передними конечностями[23]. Полезны истреблением сельскохозяйственных вредителей[24][25]:
Польза людям от жабьих ночных похождений огромна. В США попытались приблизительно оценить стоимость услуг, веками, ночь за ночью, приносимых жабами лесному и фермерскому делу: миллиарды долларов в год!Акимушкин И.И., Мир животных. Птицы, рыбы, земноводные и пресмыкающиеся. С. 391
В сумерках жабы вылезают из подвалов, нор, пещер, из-под половиц сараев и других тёмных, укромных, скрытных мест. Скачками или перебежками («жабьей рысью»), плотно прижавшись к земле, выходят эти земноводные за вредителями огородов, садов, полей и т. п.: поедают насекомых, слизней, червей[25]. Если жабу перенести из водоёма, в котором она родилась и выросла, в другой, она отправится искать свой пруд и обычно находит, если амфибию занесли не слишком далеко. Жабы возвращаются и упорно ищут воду даже в тех водоёмах, которые пересохли или засыпаны, но в которых прежде они жили головастиками[25].
Размножение

Большинству видов свойственно внешнее оплодотворение, однако некоторым — внутреннее (Nectophrynoides, Mertensophryne micranotis). Внутреннее оплодотворение встречается у четырёх (по другим исследованиям, несколько больше) родов семейства настоящих жаб[26]:
- Mertensophryne (некоторые виды)
- Nectophrynoides (предположительно, все виды)
- Вид Altiphrynoides malcolmi (один из двух видов рода Altiphrynoides)
- Nimbaphrynoides occidentalis (единственный вид в монотипическом роде Nimbaphrynoides)
Ascaphus (все виды) и Eleutherodactylus (два вида, E. coqui и E. jasperi) являются единственными другими родами лягушек, у которых есть внутреннее оплодотворение[26]. Limnonectes larvaepartus также размножаются внутренним оплодотворением[27].
Способы размножения разные — от откладывания икры в воду и последующего выхода из неё свободноплавающих личинок, до яйцеживорождения и прямого наземного развития[28]. Большинство представителей семейства нерестятся в стоячих или проточных водоёмах. Филиппинские жабы рода Pelophryne откладывают икру в пазухи листьев на высоте нескольких метров. У головастиков некоторых видов, живущих в быстротекущих ручьях (Ansonia, Atelopus) развиваются брюшные присоски, с помощью которых они прикрепляются к подводным предметам[29][12].
Типы размножения
У жаб, как и у всех бесхвостых наблюдаются два типа брачного поведения: сезонный и спонтанный. При первом типе, характерном для большинства видов, в определённые периоды года взрослые особи собираются для размножения в прудах, озёрах или ручьях. Многие жабы возвращаются к тому водоёму, в котором они развивались головастиками. В результате можно наблюдать сезонные миграции, в которые вовлечены тысячи особей. При спонтанном типе размножения взрослые жабы прибывают на места размножения, подталкиваемые некой внешней причиной (например, выпадением осадков в засушливой местности). В данном случае размножение и развитие икринок и головастиков происходит с максимальной скоростью, до того, как недолговечные водоёмы (лужи) успеют высохнуть[30].

Самцы видов жаб с сезонным типом брачного поведения обычно первыми прибывают в места размножения и остаются там продолжительное время, призывая кваканьем самок и защищая свою территорию от других самцов. Самки появляются позже лишь на время спаривания и икрометания. В этой ситуации количество самцов у кромки воды всегда превышает количество самок, поэтому среди самцов существует жёсткая конкуренция. Они взбираются на бугорок или пук травы, высунувшись наполовину из воды; крупные и сильные самцы издают более низкие звуки и захватывают самые привлекательные для размножения места. Самцы повторяют этот звук около 35-40 раз в минуту. Наблюдения показали, что самки предпочитают именно таких самцов[31]. Поскольку кваканье требует больших затрат энергии, иногда роли меняются и зовущий самец оставляет свою территорию и становится перехватчиком[30].
Многие жабы зимуют под землёй; в конце марта-апреле амфибии пробираются к воде для икрометания и спаривания. Во время дождя десятки тысяч жаб пытаются спариться. Самцы появляются первыми: они хорошо помнят свои прошлогодние пути (длиной до километра), которые ранее приводили их к водоёмам. Серые и прочие жабы запоминают также самые приметные ориентиры и могут видеть даже ультрафиолетовые лучи[25].
У видов со спонтанным типом брачного поведения основной упор делается не на выбор партнёра, а на скорость размножения. Соответственно меняется и тактика. Самец, первым обнаруживший подходящее место (лужу), начинает громко призывать сородичей, на его зов сбегаются все окрестные представители обоих полов, которые тут же подхватывают призыв, создавая хор, слышный издалека. Иногда подходящие условия отсутствуют годами, и, соответственно, эти виды не размножаются несколько лет подряд[30].
Как правило, процесс спаривания заключается в том, что самец осёдлывает самку в воде и обхватывает её туловище. Самка мечет икру, а самец покрывает её своей спермой (внешнее оплодотворение). У некоторых видов самцы могут задерживать икринки задними лапами на время, необходимое для оплодотворения[30]. В этом плане уникальными среди бесхвостых являются виды западноафриканского рода Nimbaphrynoides, которые размножаются живорождением, и виды танзанийского рода Nectophrynoides, практикующие яйцеживорождение. В обоих случаях имеет место внутреннее оплодотворение, а самка рожает полностью развитых молодых лягушек (сеголеток), которые минуют стадию головастика[32][33].
Живорождение

В Африке и в Южной Азии живут живородящие жабы Nectophrynoides длиной около шести сантиметров. Обитают в горных лесах, на земле и в траве, на листьях деревьев. Учёные узнали об этом в 1905 году, когда была открыта Nectophrynoides viviparus. У самок расширенные концы яйцеводов образовали некое подобие матки, где развиваются оплодотворенные яйца, затем зародыши. Рождаются они вполне сформированными лягушатами — до 135 молодых амфибий. Второй вид этого рода, древесная жаба Торнира, описан в 1906 году зоологом Жаном Роуксом. Рост до трех сантиметров. Новорожденных детенышей — до 35[23].
Третий вид, западная живородящая жаба, был обнаружен лишь в 1942 году за пять тысяч километров к западу от места обитания двух первых, в Гвинее. Жабы размером до 2,5 см. Ареал достаточно невелик: несколько квадратных километров травяных лугов на склонах горы Нимба. В сезон дождей, от мая до августа, этих жаб-самцов до пятисот на ста квадратных метрах луга. Затем дожди кончаются, и амфибии скрываются в разных трещинах земли почти девять месяцев, дожидаясь, пока вновь польют дожди. Перед этим самцы успевают оплодотворить самок, и те вынашивают эмбрионов. Эмбрионы кормятся, как у акул или альпийских саламандр: питательными веществами, запасенными в яйцах или зародышах, развивающихся с запозданием; каинизма у этих амфибий нет — как у червяг и скатов-хвостоколов, матка матери производит творожистую массу. Зародыши быстро растут, проглатывая её: перед рождением они, как правило, лишь втрое меньше вынашивающей их матери. Детёнышей от двух до двадцати особей, однако чем больше зародышей развивается, тем меньше ростом рождающиеся жабята[23].
Передвижение
Жабы имеют короткие задние конечности и передвигаются шагом, а не прыжками[34]. Быстрое передвижение у представителей этого семейства обеспечивают либо ускоренные движения конечностей (быстрая ходьба), либо быстрые короткие прыжки. В одном из экспериментов жаба Bufo fowleri была посажена на беговую дорожку, которая вращалась с разной скоростью. При замере количества кислорода, используемого жабой, было обнаружено, что для этого вида передвижение маленькими прыжками на длинные дистанции является энергетически неэффективным, но может быть оправдано для быстрого перемещения на короткие расстояния[35].

Жабы, обитающие в воде, адаптированы к плаванию; для строения их тела характерны сильные задние конечности и наличие плавательных межпальцевых перепонок. Перепонки увеличивают площадь поверхности ступни (наподобие ласт) и способствуют быстрому передвижению лягушек в воде. Головастики, как правило, имеют большие хвостовые плавники, придающие ускорение, направленное вперёд, когда хвост движется из стороны в сторону[23]. В воде бесхвостые наиболее беспомощны во время метаморфоза, когда хвост уже атрофирован, а ноги ещё не полностью функциональны[34].
Коммуникация
Кваканье — это звук, который производит проходящий через гортань воздух. Кваканье некоторых жаб слышно дальше чем за километр. Виды бесхвостых, обитающие преимущественно вдоль проточной воды, часто не имеют адаптаций для усиления звука, так как шумный фон делает звуковую коммуникацию неэффективной[36]. Вместо неё эти виды используют визуальные способы общения («семафорят» друг другу). Как показывают наблюдения, основная доля коммуникации приходится на привлечение самок. Самцы могут квакать в одиночестве, а могут создавать хор, когда много самцов собирается в удобных для спаривания местах. Самки предпочитают самцов, издающих более громкие и низкие звуки, которые указывают на здорового и крепкого самца, способного производить наилучшее потомство[37].
Жизненный цикл
Личинки


В яйцах развиваются личинки жаб. Через несколько недель крохотные головастики вылезают из икринок. Обычно из всех разом. Их тело, как правило, имеет овальную форму, хвост длинный, сплющенный по вертикали, приспособлен для плаванья. У головастиков хрящевой скелет, глаза лишены век, есть боковая линия, для дыхания служат жабры[38]. В начале у головастиков появляются внешние жабры, а позже внутренние (жаберный мешок закрывает жабры и передние ноги). Развивающиеся лёгкие служат дополнительным дыхательным органом. У головастиков нет настоящих зубов, но у многих видов челюсти покрыты параллельными рядами кератиновых зубчиков (два ряда на верхней челюсти, три ряда на нижней и роговой клюв). Количество рядов и точная морфология рта варьируют у разных видов и могут служить диагностическим признаком[39]. Плавают личинки многотысячными стайками. Живыми лентами в метр шириной и несколько метров длиной вьются они в пруду. То у поверхности, то погружаются на дно. Все личинки разом совершают манёвры: плывут в одну сторону и в одном ритме[40].
Метаморфоз

Через несколько месяцев начинается метаморфоз, во время которого системы организма резко перестраиваются во взрослую форму. Как правило, метаморфоз длится около суток. Он начинается с выработки гормона тироксина, который влияет на развитие тканей. Так, изменение органов дыхания включает в себя развитие лёгких параллельно с исчезновением жабр и жаберного мешка. Передние конечности становятся видимыми. Нижняя челюсть приобретает характерную для хищника форму, кишечник укорачивается. Нервная система адаптируется под стереоскопическое зрение и слух, а также под новые способы передвижения и питания. Глаза сдвигаются выше, формируются веки и связанные с ними железы. Модифицируются органы слуха (появляется слуховая перепонка и среднее ухо). Кожа становится толще и крепче, боковая линия исчезает (у большинства видов), формируются кожные железы. На последнем этапе метаморфоза исчезает хвост, ткани которого идут на развитие конечностей[41][42]. Постепенно жабята выходят на сушу. Заканчивается метаморфоз практически одновременно у всех головастиков[40].
Взрослые особи
В неволе зарегистрирована продолжительность жизни представителей семейства до 40 лет, этого возраста достигла обыкновенная жаба. Жаба-ага жила в неволе 24 года, жаба Бломберга — 28 лет, колорадская жаба — до 15 лет, а Peltophryne peltocephala fustiger — 20 лет[43].
Распространение
Распространены повсеместно, за исключением Арктики, Антарктики, Гренландии, Мадагаскара, Новой Зеландии и многих островов Тихого океана[44]. В Австралии и Гвинее обитает ввезённая из Америки жаба-ага[45].
На территории России распространены лишь представители рода Bufo, всего 6 видов: обыкновенная, кавказская, дальневосточная, зелёная, камышовая и монгольская жабы[13].
Естественные враги и паразиты
На жаб охотятся многие хищники, в том числе цапли, ястребы, рыбы, крупные саламандры, змеи (в том числе ужи, полозы), черепахи, более крупные лягушки, ежи, сороки, вороны, хищные жуки и сороконожки, еноты, скунсы, хорьки и другие. Опасны люди, посыпающие поля инсектицидами и ДДТ. В дельте Миссисипи эти химикаты практически полностью истребили, уничтожили обитающих там жаб; немногие уцелели и сумели приспособиться к ядовитым веществам[23].

Опасны также для жаб лягушкоедки, или люцилии. Лягушкоедка — это зелёная падальная муха, паразитирующая на лягушках и жабах. В период размножения летом мухи откладывают свои белые продолговатые яйца на спину или голову жаб и других амфибий. Хозяевами паразита являются: обыкновенная, камышовая, зелёная жабы[46]. Вылупившиеся личинки проникают через носовые отверстия в голову хозяина, где въедаются в мягкие ткани ещё живущего животного. Через несколько дней передняя часть морды и головы может полностью деформироваться и исказиться. Когда личинки добираются до мозга или других жизненно важных органов, функции организма нарушаются и амфибия погибает. В своей последней фазе личинки почти полностью съедают труп животного, включая кожу и соединительные ткани, при определённых обстоятельствах — даже хрящи и кости[47].
Количество мутаций и генетических дефектов, наблюдаемых у жаб, как и у всех бесхвостых, возросло за время наблюдения с 1990-х по 2003 год. Одним из распространённых дефектов являются недостающие или лишние конечности. Различные гипотезы по поводу причин, вызывающих эти дефекты, включают в себя увеличение ультрафиолетового излучения, попадающего на икру, загрязнение сельскохозяйственными ядохимикатами и паразитические заболевания, как, например, заражение трематодами Ribeiroia ondatrae. Возможно, что все эти факторы действуют вместе (радиационный и химический стресс понижает сопротивление организмов паразитам). Дефекты конечностей ухудшают подвижность и, как следствие, шансы животного дожить до половой зрелости[48][49].
Самки комаров рода Corethrella питаются кровью бесхвостых амфибий, в том числе и жаб. Жертву они находят, ориентируясь по звуковым брачным сигналам. Самцы жаб издают сложные и простые позывные сигналы. Сложные сигналы предпочитаются, как самками комаров, так и самками лягушек. Если самцы издают простые звуки, они меньше подвергаются нападению кровососов, но и вероятность спаривания таких самцов уменьшается. Комары служат переносчиками трипаносомозов лягушек[50].
Специализированными хищниками на многих жаб являются жуки и их личинки рода Epomis в составе семейства жужелиц[51].
Угрозы и охрана

Исследования, начатые в 1950-х годах, указывают на значительное сокращение численности многих видов бесхвостых, в том числе и жаб. Среди этих видов числится и оранжевая жаба из Коста Рики. Её исчезновение вызвало серьёзную озабоченность исследователей, связанную с тем, что эта жаба обитала в заповеднике «Monteverde Cloud Forest Reserve» и её вымирание не могло быть вызвано напрямую человеческим вмешательством. В других местах снижение численности обусловлено разрушением среды обитания, поллютантами, климатическими изменениями и внедрением чужеродных хищников, паразитов и конкурентов. Особенно разрушительными для популяций жаб и вообще всех бесхвостых считаются инфекционные заболевания хитридиомикоз и ранавирус[52][53].
В культуре
В европейском фольклоре жабы ассоциировались с ведьмами как фамилиары, источники ингредиентов для магических зелий, а иногда имели собственные магические свойства[54]. В иконографии жаба является атрибутом смерти или алчности. Кроме того, жаба, пожирающая гениталии обнаженной женской фигуры, символизирует похоть[55]. Легенда Панамы гласит, что удача сопутствует тому, кто увидит панамскую золотую лягушку (Atelopus zeteki). В варианте этой легенды говорится, что, умирая, эти лягушки превращаются в золотой талисман уаку[56]. В мультфильмах «Шрек 2» и «Шрек 3» жаба — истинный облик короля Гарольда.
Классификация
На февраль 2023 года в семейство включают 52 рода и 642 видов[44]:
- Adenomus Cope, 1861 (2 вида)
- Altiphrynoides Dubois, 1987 (2 вида)
- Amazophrynella Fouquet et al., 2012 (13 видов)
- Anaxyrus Tschudi, 1845 (25 вида)
- Ansonia Stoliczka, 1870 — Анзонии (38 вида)
- Atelopus Dumeril & Bibron, 1841 — Ателопы, или арлекины (99 видов)
- Barbarophryne Beukema et al., 2013 (1 вид)
- Barbarophryne brongersmai (Hoogmoed, 1972) — Марокканская жаба
- Beduka Dubois, Ohler, and Pyron, 2021 (2 вида)
- Beduka amboli Dubois, Ohler, and Pyron, 2021
- Beduka koynayensis (Soman, 1963) — Деканская жаба
- Blaira Dubois, Ohler, and Pyron, 2021 (2 вида)
- Blaira ornata (Gunther, 1876) — Украшенная анзония
- Blaira rubigina (Pillai and Pattabiraman, 1981) — Пятнистобокая анзония
- Blythophryne Chandramouli et al., 2016 (1 вид)
- Blythophryne beryet Chandramouli et al., 2016
- Bufoides Pillai & Yazdani, 1973 — Скальные жабы (2 вида)
- Bufoides kempi (Boulenger, 1919) — Ассамская жаба
- Bufoides meghalayanus (Yazdani & Chanda, 1971) — Скальная жаба
- Bufotes Rafinesque, 1814 — Зелёные жабы (13 видов)
- Capensibufo Grandison, 1980 — Капские жабы (5 видов)
- Churamiti Channing & Stanley, 2002 (1 вид)
- Churamiti maridadi Channing & Stanley, 2002
- Dendrophryniscus Jimenez de la Espada, 1870 — Древесные арлекины (16 видов)
- Didynamipus Andersson, 1903 — Четырёхпалые жабы (1 вид)
- Didynamipus sjostedti Andersson, 1903 — Четырёхпалая жаба
- Duttaphrynus Frost et al., 2006 (28 видов)
- Epidalea Cope, 1864 (1 вид)
- Epidalea calamita Laurenti, 1768 — Камышовая жаба
- Frostius Cannatella, 1986 (2 вида)
- Frostius erythrophthalmus Pimenta & Caramaschi, 2007
- Frostius pernambucensis (Bokermann, 1962) — Пернамбукский арлекин
- Incilius Cope, 1863 (39 видов)
- Ingerophrynus Frost et al., 2006 (12 видов)
- Laurentophryne Tihen, 1960 — Лорановы жабы (1 вид)
- Laurentophryne parkeri (Laurent, 1950) — Лоранова жаба
- Leptophryne Fitzinger, 1843 — Тонкотелые жабы (3 вида)
- Leptophryne borbonica (Tschudi, 1838)
- Leptophryne cruentata (Tschudi, 1838) — Кровяная жаба
- Leptophryne javanica Hamidy et al., 2018
- Melanophryniscus Gallardo, 1961 — Черноспинки (31 вид)
- Mertensophryne Tihen, 1960 — Мертенсовы жабы (14 видов)
- Metaphryniscus Senaris et al., 1994 (1 вид)
- Metaphryniscus sosai Senaris, Ayarzaguena & Gorzula, 1994
- Nannophryne Gunther, 1870 (4 вида)
- Nannophryne apolobambica (De la Riva, Rios & Aparicio, 2005)
- Nannophryne cophotis (Boulenger, 1900) — Горная жаба
- Nannophryne corynetes (Duellman & Ochoa-M., 1991)
- Nannophryne variegata Gunther, 1870 — Патагонская жаба
- Nectophryne Buchholz & Peters, 1875 — Лазающие жабы (2 вида)
- Nectophrynoides Noble, 1926 — Живородящие жабы (13 видов)
- Nimbaphrynoides Dubois, 1987 (1 вид)
- Oreophrynella Boulenger, 1895 — Гвианские арлекины (9 видов)
- Osornophryne Ruiz-Carranza & Hernandez-Camacho, 1976 — Тучные арлекины (11 видов)
- Parapelophryne Fei, Ye & Jiang, 2003 (1 вид)
- Parapelophryne scalpta (Liu at al., 1973) — Хайнаньская жабка
- Pedostibes Gunther, 1876 — Древесные жабы (1 вид)
- Pelophryne Barbour, 1938 — Жабки (13 видов)
- Peltophryne Fitzinger, 1843 — Карибские жабы (14 видов)
- Phrynoidis Fitzinger, 1842 (2 вида)
- Phrynoidis asper (Gravenhorst, 1829) — Шероховатая жаба
- Phrynoidis juxtaspera (Inger, 1964) — Исполинская жаба
- Poyntonophrynus Frost et al., 2006 (11 видов)
- Pseudobufo Tschudi, 1838 — Водяные жабы (1 вид)
- Rentapia Chan at al., 2016 (3 вида)
- Rhaebo Cope, 1862 (14 видов)
- Rhinella Fitzinger, 1826 (92 вида)
- Sabahphrynus Matsui, Yambun & Sudin, 2007 (1 вид)
- Sabahphrynus maculatus (Mocquard, 1890) — Пятнистая жаба
- Schismaderma Smith, 1849 — Красные жабы (2 вида)
- Schismaderma branchi Baptista, Vaz Pinto, Keates, Edwards, Rodel, and Conradie, 2021
- Schismaderma carens (Smith, 1848) — Красная жаба
- Sclerophrys Tschudi, 1838 (45 видов)
- Sigalegalephrynus Smart et al., 2017 (5 видов)
- Sigalegalephrynus burnitelongensis Sarker at al., 2019
- Sigalegalephrynus gayoluesensis Sarker at al., 2019
- Sigalegalephrynus harveyi Sarker at al., 2019
- Sigalegalephrynus mandailinguensis Smart at al., 2017
- Sigalegalephrynus minangkabauensis Smart at al., 2017
- Strauchbufo Fei, Ye & Jiang, 2012 (1 вид)
- Truebella Graybeal & Cannatella, 1995 (2 вида)
- Truebella skoptes Graybeal & Cannatella, 1995
- Truebella tothastes Graybeal & Cannatella, 1995
- Vandijkophrynus Frost et al., 2006 (6 видов)
- Werneria Poche, 1903 — Вернерии (6 видов)
- Wolterstorffina Mertens, 1939 — Вольтерсторфовы жабы (3 вида)
Виды incertae sedis
- Bufo scorteccii Balletto and Cherchi, 1970 — Йеменская жаба
Галерея
-

-

Амазонский арлекин
-

Карибская жаба
-





Примечания
- ↑ 1 2 Источник русских названий: Ананьева Н. Б., Боркин Л. Я., Даревский И. С., Орлов Н. Л. Пятиязычный словарь названий животных. Амфибии и рептилии. Латинский, русский, английский, немецкий, французский. / под общ. ред. акад. В. Е. Соколова. — М.: Рус. яз., 1988. — С. 35. — 10 500 экз. — ISBN 5-200-00232-X.
- ↑ Фасмер М. Жаба // Этимологический словарь русского языка.
- ↑ Винокур Г. О., проф. Ларин Б. А., Ожегов С. И., Томашевский Б. В., проф. Ушаков Д. Н. Толковый словарь русского языка: В 4 т / Под ред. Ушакова Д. Н. — М.: Государственный институт «Советская энциклопедия»; ОГИЗ (т. 1); Государственное издательство иностранных и национальных словарей (т. 2-4), 1935—1940. — 45 000 экз.
- ↑ Ананьева Н. Б., Боркин Л. Я., Даревский И. С., Орлов Н. Л. Пятиязычный словарь названий животных. Амфибии и рептилии. Латинский, русский, английский, немецкий, французский. / под общ. ред. акад. В. Е. Соколова. — М.: Рус. яз., 1988. — 560 с. — 10 500 экз. — ISBN 5-200-00232-X.
- ↑ Jennifer B. Pramuk, Tasia Robertson, Jack W. Sites, Brice P. Noonan. Around the world in 10 million years: biogeography of the nearly cosmopolitan true toads (Anura: Bufonidae) (англ.) // Global Ecology and Biogeography. — 2007-08-16. — Vol. 0, iss. 0. — P. 070817112457001–???. — ISSN 1466-8238 1466-822X, 1466-8238. — doi:10.1111/j.1466-8238.2007.00348.x. Архивировано 27 октября 2022 года.
- ↑ 1 2 3 Philippe J. R. Kok, Sebastian Ratz, Ross D. MacCulloch, Amy Lathrop, Raheleh Dezfoulian. Historical biogeography of the palaeoendemic toad genus Oreophrynella (Amphibia: Bufonidae) sheds a new light on the origin of the Pantepui endemic terrestrial biota (англ.) // Journal of Biogeography. — 2018-01. — Vol. 45, iss. 1. — P. 26–36. — doi:10.1111/jbi.13093. Архивировано 27 октября 2022 года.
- ↑ Daniel M. Portik, Theodore J. Papenfuss. Historical biogeography resolves the origins of endemic Arabian toad lineages (Anura: Bufonidae): Evidence for ancient vicariance and dispersal events with the Horn of Africa and South Asia (англ.) // BMC Evolutionary Biology. — 2015-12. — Vol. 15, iss. 1. — P. 152. — ISSN 1471-2148. — doi:10.1186/s12862-015-0417-y. Архивировано 8 ноября 2020 года.
- ↑ Kin Onn Chan, L. Lee Grismer, Anil Zachariah, Rafe M. Brown, Robin Kurian Abraham. Polyphyly of Asian Tree Toads, Genus Pedostibes Günther, 1876 (Anura: Bufonidae), and the Description of a New Genus from Southeast Asia (англ.) // PLOS ONE / Helge Thorsten Lumbsch. — 2016-01-20. — Vol. 11, iss. 1. — P. e0145903. — ISSN 1932-6203. — doi:10.1371/journal.pone.0145903.
- ↑ S. R. Chandramouli, Karthikeyan Vasudevan, S Harikrishnan, Sushil Kumar Dutta, S Jegath Janani. A new genus and species of arboreal toad with phytotelmonous larvae, from the Andaman Islands, India (Lissamphibia, Anura, Bufonidae) // ZooKeys. — 2016-01-20. — Т. 555. — С. 57–90. — ISSN 1313-2989 1313-2970, 1313-2989. — doi:10.3897/zookeys.555.6522. Архивировано 9 октября 2022 года.
- ↑ G.A. Boulenger. LXI.— Description of a new Batrachian (Oreophryne Quelchii) discovered by Messrs. J. J. Quelch and F. McConnell on the summit of Mount Roraima (англ.) // Annals and Magazine of Natural History. — 1895-06. — Vol. 15, iss. 90. — P. 521–522. — ISSN 0374-5481. — doi:10.1080/00222939508680212.
- ↑ The Guinness book of records 1998.. — Enfield: Guinness, 1997. — С. 249. — 352 pages с. — ISBN 0-85112-044-X, 978-0-85112-044-7.
- ↑ 1 2 Gray, 1825 : A synopsis of the genera of reptiles and Amphibia, with a description of some new species. Annals of Philosophy, London, ser.2, vol.10, p.193–217 [1].
- ↑ 1 2 Ананьева Н. Б., Боркин Л. Я., Даревский И. С., Орлов Н. Л. Энциклопедия природы России. Земноводные и пресмыкающиеся / ред. серии д. б. н. Минин А. А. — М.: ABF, 1998. — С. 96. — 10 000 экз. — ISBN 5-87484-041-9.
- ↑ Земноводные. Пресмыкающиеся: Энциклопедический справочник. - Мн., БелЭн, 1996. ISBN 985-11-0067-6
- ↑ Кузьмин С. Л. Земноводные бывшего СССР. — М.: Товарищество научных изданий КМК, 2012. — 2-е изд. — С. 129. — 370 с. — ISBN 978-5-87317-871-1
- ↑ Zweifel, Richard G. Encyclopedia of Reptiles and Amphibians / Cogger, H.G. ; Zweifel, R.G.. — San Diego : Academic Press, 1998. — P. 91–92. — ISBN 978-0-12-178560-4.
- ↑ Halliday, Tim R., and Kraig Adler (editors). The New Encyclopedia of Reptiles & Amphibians. Facts on File, New York, 2002.
- ↑ African Amphibians Bufonidae. The goal of this website is to develop an authoritative, community-driven resource for information on African amphibians (2017). Дата обращения: 16 сентября 2017. Архивировано 16 сентября 2017 года.
- ↑ Vitt, L. J., & Caldwell, J. P. (2014). Herpetology: an introductory biology of amphibians and reptiles. Academic Press.
- ↑ Brown, Federico D.; Del Pino, Eugenia M.; Krohne, Georg. Bidder's organ in the toad Bufo marinus: Effects of orchidectomy on the morphology and expression of lamina-associated polypeptide 2 (англ.) // Development, Growth & Differentiation : journal. — 2002. — December (vol. 44, no. 6). — P. 527—535. — ISSN 1440-169X. — doi:10.1046/j.1440-169X.2002.00665.x. Архивировано 16 сентября 2017 года.
- ↑ Piprek, Rafal P., et al. «Bidder’s Organ — Structure, Development and Function.» The International Journal of Developmental Biology, vol. 58, no. 10-11-12, 2014, pp. 819-27. Crossref, doi:10.1387/ijdb.140147rp.
- ↑ Brown, Federico D.; Del Pino, Eugenia M.; Krohne, Georg (December 2002). “Bidder's organ in the toad Bufo marinus: Effects of orchidectomy on the morphology and expression of lamina-associated polypeptide 2”. Development, Growth & Differentiation [англ.]. 44 (6): 527—535. DOI:10.1046/j.1440-169X.2002.00665.x. ISSN 1440-169X. PMID 12492511. S2CID 44753338.
- ↑ 1 2 3 4 5 Акимушкин, 1989, с. 393, 394.
- ↑ Книпович Н. М.,. Жабы // Энциклопедический словарь Брокгауза и Ефрона : в 86 т. (82 т. и 4 доп.). — СПб., 1890—1907.
- ↑ 1 2 3 4 Акимушкин, 1989, с. 391.
- ↑ 1 2 Vitt, Laurie J. Herpetology: An Introductory Biology of Amphibians and Reptiles / Laurie J. Vitt, Janalee P. Caldwell. — 4th. — Academic Press, 2014. — P. 122.
- ↑ Iskandar, D. T.; Evans, B. J.; McGuire, J. A. (2014). “A novel reproductive mode in frogs: a new species of fanged frog with internal fertilization and birth of tadpoles”. PLOS ONE. 9 (12): e115884. Bibcode:2014PLoSO...9k5884I. DOI:10.1371/journal.pone.0115884. PMC 4281041. PMID 25551466.
- ↑ Zweifel, Richard G. Encyclopedia of Reptiles and Amphibians (неопр.) / Cogger, H.G.; Zweifel, R.G.. — San Diego: Academic Press, 1998. — С. 91—92. — ISBN 0-12-178560-2.
- ↑ Stebbins, Robert. Western Reptiles & Amphibians (3rd ed.). Houghton Mifflin Co., 2003.
- ↑ 1 2 3 4 Stebbins, Robert C.; Cohen, Nathan W. A Natural History of Amphibians (англ.). — Princeton University Press, 1995. — P. 154‑162. — ISBN 0-691-03281-5.
- ↑ Davies, N. B.; Halliday, T. R. Deep croaks and fighting assessment in toads Bufo bufo (англ.) // Nature : journal. — 1978. — Vol. 274. — P. 683‑685. — doi:10.1038/274683a0.
- ↑ Channing, Alan; Howell, Kim M. Amphibians of East Africa (англ.). — Comstock Publishing, 2006. — P. 104‑117. — ISBN 0801443741.
- ↑ Sandberger, L.; Hillers, A.; Doumbia, J.; Loua, N-S.; Brede C.; Rödel, M-O. Rediscovery of the Liberian Nimba toad, Nimbaphrynoides liberiensis (Xavier, 1978) (Amphibia: Anura: Bufonidae), and reassessment of its taxonomic status (англ.) // Zootaxa : journal. — Magnolia Press, 2010. — Vol. 4355. — P. 56‑68. — ISSN 1175-5334. Архивировано 29 октября 2013 года.
- ↑ 1 2 Anura. Encyclopædia Britannica Online. Дата обращения: 12 июня 2012. Архивировано 17 августа 2012 года.
- ↑ Walton, M.; Anderson, B. D. The aerobic cost of saltatory locomotion in the fowler's toad (Bufo woodhousei fowleri) (англ.) // The Journal of Experimental Biology : journal. — The Company of Biologists, 1988. — Vol. 136. — P. 273‑288. Архивировано 22 декабря 2015 года.
- ↑ Nafis, Gary. Ascaphus truei: Coastal Tailed Frog. California Herps (2012). Дата обращения: 1 августа 2012. Архивировано 5 августа 2012 года.
- ↑ Gerhardt, H. C. The evolution of vocalization in frogs and toads (англ.) // Annual Review of Ecology, Evolution, and Systematics. — Annual Reviews, 1994. — Vol. 25. — P. 293‑324. — doi:10.1146/annurev.es.25.110194.001453.
- ↑ Duellman, W. E.; Zug, G. R. Anura: From tadpole to adult. Encyclopædia Britannica. Дата обращения: 13 июля 2012. Архивировано 17 октября 2012 года.
- ↑ Stebbins, Robert C.; Cohen, Nathan W. A Natural History of Amphibians (англ.). — Princeton University Press, 1995. — P. 179‑194. — ISBN 0-691-03281-5.
- ↑ 1 2 Акимушкин, 1989, с. 392.
- ↑ Balinsky, Boris Ivan. Animal development: Metamorphosis. Encyclopedia Britannica. Дата обращения: 10 августа 2012. Архивировано 17 октября 2012 года.
- ↑ Campbell, Neil A.; Reece, Jane B. Biology (Sixth edition) (англ.). — Pearson Education, 2002. — P. 692. — ISBN 0-201-75054-6.
- ↑ Slavens, Frank; Slavens, Kate. Frog and toad index. Longevity. Дата обращения: 4 июля 2012. Архивировано 17 октября 2012 года.
- ↑ 1 2 Frost, Darrel R. Bufonidae. Amphibian Species of the World: an Online Reference. Version 6.0. American Museum of Natural History (2014). Дата обращения: 22 января 2015. Архивировано 28 января 2015 года.
- ↑ Amphibia Web. Bufonidae. Provides information on amphibian declines, natural history, conservation, and taxonomy (2017). Дата обращения: 16 сентября 2017. Архивировано 30 марта 2015 года.
- ↑ V. Neumann & F. Meyer: Lucilia bufonivora MONIEZ, 1876 — ein euryxener Amphibienparasit. — Mitteilungen aus dem Zoologischen Museum Berlin 70 (1994): 331—341.
- ↑ Vít Zavadil: Zum Parasitismus der Krötengoldfliege (Lucilia bufonivora MONIEZ, 1876) auf Erdkröten (Bufo bufo) — Abwehrverhalten und limitierende Faktoren. — Zeitschrift für Feldherpetologie 4 (1997): 1—12.
- ↑ Blaustein, Andrew R.; Johnson, Pieter T. J. The complexity of deformed amphibians (англ.) // Frontiers in Ecology and the Environment. — Ecological Society of America, 2003. — Vol. 1, no. 2. — P. 87—94. Архивировано 29 октября 2013 года.Архивированная копия. Дата обращения: 27 августа 2012. Архивировано из оригинала 29 октября 2013 года.
- ↑ Burkhart J. G., Ankley G., Bell H., Carpenter H., Fort D., Gardiner D., Gardner H., Hale R., Helgen J. C., Jepson P., Johnson D., Lannoo M., Lee D., Lary J., Levey R., Magner J., Meteyer C., Shelby M. D., Lucier G. Strategies for assessing the implications of malformed frogs for environmental health (англ.) // Environmental Health Perspectives : journal. — 2000. — Vol. 108, no. 1. — P. 83—90. — doi:10.2307/3454299. — PMID 10620528. — . — PMC 1637865.
- ↑ Acoustic preferences and localization performance of blood-sucking flies (Corethrella Coquillett) to túngara frog calls. academic.oup.com. Дата обращения: 12 сентября 2022. Архивировано 2 сентября 2018 года.
- ↑ Gil Wizen, Avital Gasith. An Unprecedented Role Reversal: Ground Beetle Larvae (Coleoptera: Carabidae) Lure Amphibians and Prey upon Them (англ.) // PLOS ONE. — 21 сент. 2011 г.. — Vol. 6, iss. 9. — P. e25161. — ISSN 1932-6203. — doi:10.1371/journal.pone.0025161. Архивировано 2 ноября 2022 года.
- ↑ Voordouw M. J., Adama D., Houston B., Govindarajulu P. Prevalence of the pathogenic chytrid fungus, Batrachochytrium dendrobatidis, in an endangered population of northern leopard frogs, Rana Pipiens (англ.) // BMC Ecology : journal. — 2010. — Vol. 10, no. 6. — doi:10.1186/1472-6785-10-6.
- ↑ Harp, Elizabeth M.; Petranka, James W. Ranavirus in wood frogs (Rana sylvatica): Potential sources of transmission within and between ponds (англ.) // Journal of Wildlife Diseases : journal. — 2006. — Vol. 42, no. 2. — P. 307—318. — PMID 16870853.
- ↑ Burns, William E. Witch Hunts in Europe and America: An Encyclopedia (англ.). — Greenwood Publishing Group, 2003. — P. 7. — ISBN 0-313-32142-6.
- ↑ Hall, J. (2000). Hall's Iconografisch Handboek. Leiden: Primavera Pers. ISBN 9074310052
- ↑ Gratwicke, B. The Panamanian Golden Frog. Panama Amphibian Rescue and Conservation Project (2009). Архивировано 17 октября 2012 года.
Литература
- Акимушкин И. И. Мир животных. Птицы, рыбы, земноводные и пресмыкающиеся. — Мысль, 1989. — 466 с.
- Amphibian Species of the World 5.1 - Bufonidae. Дата обращения: 10 апреля 2008. Архивировано из оригинала 15 июля 2012 года.
- Stebbins R.C.. Western Reptiles & Amphibians (3rd ed.). Houghton Mifflin Co., 2003.
- Halliday, Tim R., and Kraig Adler (editors). The New Encyclopedia of Reptiles & Amphibians. Facts on File, New York, 2002.
Ссылки
- Tolweb.org: Bufonidae
- Bufonidae.com
- Amphibian and Reptiles of Peninsular Malaysia — Family Bufonidae
- FED.us
- Bufonidae recordings из British Library Sound Archive
| Таксономия | ||||
|---|---|---|---|---|
