ජාන
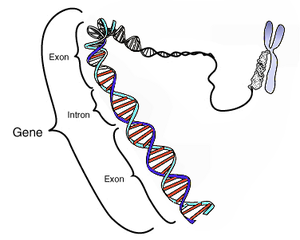

DNA ද්විත්ව හෙලික්සයේ භෂ්ම 4 සහිත රසායනික ව්යුහය බහුතරයක් ජිව සංවිධාන ඔවුන්ගේ ජාන රඳවා ගන්නේ DNA දිගු දාමවලයි. DNA වල ඇති දාමය භෂ්ම වර්ග 4කින් යුතු නියුක්ලියෝටයිඩ වර්ග 4කින් යුතුයි. ඒවා උප ඒකකයි. එනම් ඇඩිනින් සයිටොසීන්, ගුඇනින්, තයිමීන්. සෑම නියුක්ලියෝටයිඩ උප ඒකකයක්ම සංඝටක 3කින් යුතුයි. පොස්ෆේට කාණ්ඩය, ඩිඔක්සිරයිබෝස් සීනි වළල්ල නියුක්ලියෝ භෂ්ම එම නිසා DNA හෝ RNA වල ඇති නියුක්ලියෝටයිඩ දර්ශීයව භෂ්ම නමින් හඳුන්වන අතර, ඒවායේ ඇති පියුරීන්, පිරිමිඩින් නියම භෂ්ම සංඝටක වන ඇඩ්නින්, සයිටොඩින්, ගඇනින්, තයිමීන්වලට අනුව හඳුන්වයි. බහුතරය DNA ව්යුහය වන්නේ ද්විත්ව හෙලික්සයයි. DNA අණු 2ක දක්ෂිණාවර්ථව සර්පිලාකාරව එක වටා එකක් ලෙස ඇඹරී මෙය තනයි. මෙම ව්යුහයට අනුව භෂ්ම ගැලපීමේ රීතියට අනුව ගුඇනින් සයිටොසීන් සමඟත් ඇඩිනින් තයිමීන් සමඟත් යුගල විය යුතුයි. (එක් යුගලයක්1 පියුරිනයක් හා 1 පිරිමීඩිනයකින් යුක්තයි) මෙම යුගල වන ගුඇනින්, සයිටොසින් අතර H බන්ධන 3ක් ද ඇඩිනින් තයිමීන් අතර H බන්ධන 2ක් ද ඇත. එම නිසා DNA ද්විත්ව හෙලික්සයේ DNA අණු 2, එකකට එකක් අනුරූප වේ. පෙන්ටෝස් සීනි අණුවෙහි රසායනික සංයුක්තය නිසා DNA රැහැනට දිශාගත වීම වේ. DNA මහාණුවේ එක් කොටසක විවෘත වූ හයිඩොසිල් කාණ්ඩයක් ඩිඔක්සිරයිබෝස් මත තිබෙන අතර මෙය අණුවේ 31 අන්තය නම් වේ. අනෙක් අන්තයේ විවෘත පොස්පේට් කාණ්ඩයකි. මෙය 51 අන්තයයි. බොහෝ සෛල ක්රියාකාරකම්වලට මෙම DNA වල දිශාගත වීම වැදගත් වේ. DNA ද්විත්ව හෙලික්සයන් සෑම විටම දිශාවක් ඇති නිසා (එක් රැහැන් 51-31 ද යුගල වූ අනික් රැහැන් 31-51 දිශාවට සිදු වේ.හේතුව අලුත් කුඩා අණු විවෘත වූ 31 හයිඩ්රොක්සිල් කොන න්යෂ්ටික අම්ල ප්රිය කරන ස්ථානයක් nucleophile ලෙස ක්රියා කර එහි දී ජල විච්ඡේදක ප්රතික්රියාවකින් එකතු වේ. DNA වල ඇතුළු වූ 1 සංයුක්ත වූ ජාන ප්රකාශනය ඇරඹෙන්නේ ජාන RNA වලට පිටපත් වීමෙනි. RNA තව නියුක්ලික් අම්ල ආකාරයක් වන අතර DNA වලට සමානය නමුත් ඒවායේ ඒකකවල අඩංගු වන්නේ රයිබෝස් සීනි අණුවකි. (ඩිඔක්සිරයිබෝස් නොවේ) තවද තයිමින් වෙනුවට යුරැසිල භෂ්මයක් ඇත. DNA වලට වඩා RNA වල ස්ථාවරභාවය අඩු අතර දර්ශීයව තනි රැහැනකින් යුතුයි. ප්රෝටීනවලට කේත සහිත ජාන කෝඩෝන නම් නියුක්ලියෝටයිඩ අනුපිළිවෙලවල් 3කින් යුතු කොටස් අඩංගුය. එම කෝඩෝන ජාන “භාෂාවේ” “වචන” ලෙස ක්රියා කරයි. ප්රෝටීන පරිවර්තනයේ දී කෝඩාන හා ඇමීනෝ අම්ල අතර ඇති අනුරූපතාව ප්රවේණි කේතයන් සංඥා විශේෂිත වේ. සෑම දන්නා ජීවී සංවිධානයකට ප්රවේණික කේතය එකම වේ.
හැඳින්වීම
ජාන යනු ප්රවේණික අංශයට අනුරූප ජානමය අනුක්රමයේ නිශ්වසනය කළ කොටසක් වේ. මෙය යාමක කොට්ඨාසය අනුවත් කොට්ඨාස හා/හෝ අනෙකුත් කෘක්යාත්මක අනුක්රමිකතා කොට්ඨාසයන්හි හා කටයුතු කරයි. ජීවින්ගේ අවයවයන්හි වර්ධනය සහ රූපානුදර්ශනය ජාන එකිනෙක හා සම්බන්ධව කටයුතු තිරිමෙන් හෝ අවට පරිසර සම්බන්ධව කටයුතු කිරිමෙන් හෝ සිදු කරනු ඇත.
සාමාන්ය ව්යවහාරයේදි ජාන යන පදය පවුල් පරම්පරාවෙන් උරුම වු ගති ලක්ෂණ පිළිබද විස්තර කිරිමේ දි උපයෝගි කර ගනී. උදාහරණයක් වශයෙන් උස ජාන හෝ නරක ජාන දැක්විය හැකිය. මෙය බොහෝ විට රූපානු දර්ශන ආකාරයෙන් දක්වා ඇත. මනුත් මෙම ජනා හැදින්විම සදහා භාවිතා කළ යුතු නිවැරදි විද්යාත්මක නාමය වන්නේ ඇලීලය යවේ.
සෛලයක ජාන සතුව ඉතා දිගු DNA තන්තුවක් පවතින අතර එම DNA තන්තුවක සතුව ජානයේ ක්රියාකාරිත්වය පාලනය කරන පාලකයන් ඇත. කෙන්ති කරණ අනුපිළිවෙනින් ජානයක අනුපිළිවෙල සිදු නොවන විට පිටපත් කිරිම පෙස හැදින්වන ක්රියාවලියකට ජාන භාජනය වන අතර එහිදි ජානයේ තොරතුරු සහිත RNA පිටපතක් නිර්මාණය වේ. ජානය කේතය ඔස්සේ මෙම RNA හට ප්රොටීයන්හි සංකලනය හැසිරවිය හැකිය. නමුත් සමහර අවස්ථාවන්හිදි RNA සෘජුවම භාවිතා කරනු ලැබේ.
ජීවියෙකු සතු ජාන සමුහය ජෙනෝම් ලෙසින් හදුන්වනු ලබයි. ඒක සෛලික ශුද්ර ජීවියෙකු වන ඇමීබාවා සතුව ලොව පවතින වැඩිම නෛරම් ප්රමාණයක් අයිති වන අතර අබීබවා සතුව මනුෂ්යායට වඩා 200 ගුණයකට වඩා වැඩි ජාන ප්රමාණයක් අයිති වේ.
සමහර අවස්ථාවලදී RNA ,ජාන වලින් ප්රෝටීන නිෂ්පාදනය කිරීමේදී සිදුකරන අතරමැදි නිෂ්පාදනයකි. කෙසේ වුවද අනෙක් ජාන අනුපිලිවෙලවලට RNA අණු ඇත්නම් ක්රියාකාරී නිෂ්පාදනය. උදා- රයිබොසෙම නම් වූ RNA වලට එන්සයිම ක්රියා කරන අතර mRNA නියාමක කෘත්යක් සිදුකරයි. මෙම RNA අනුපිලිවෙලින් නිෂ්පාදනය වන RNA,RNA ජාන නමි වේ. සමහර වෛරස ඔවුන්ගේ සම්පූර්ණ ජිනෝමයම RNA ලෙස ගබඩාකරන අතර DNA අඩංගු නොවේ. ඔවුන් ජාන ඔවුන් ජාන ගබඩා කිරීමට RNA යොදා ගන්නා නිසා ඔවුන්ගේ සෛලික ධාරකයන් විසින් ආසාධනය වූ විගස ඔවුන්ගේ ප්රෝටීන නිශ්පාදනය කරන අතර පිටපත් කිරීමටද කාලයක්ද වැය නොවේ. අනෙක් අතින් RNA රෙට්රොවයිරස (HIV වැනි) ප්රතිපක්ෂ පිටපත් කිරීම මගින් RNA තුල ඇති ජිනෝමය ප්රෝටීන නිෂ්පාදනයට පෙර RNA බවට පත්කරගනී .2006 දී ප්රංශ විද්යාඥයින් පැනයට විසඳුම් සොයාගනී. උදා - ලෙස මීයන්ගේ RNA මධ්ය වූ ප්රවේනිය kit නම් ජානයේ කෘත්ය අහිමි වූ විකෘතිය නිසා සුදු වලිගා මීයන්ට ඇතිවේ. ඔවුන්ගේ පැටවුන්ට kit ජානය තිබුනද හිමිවන්නේ සුදු වලිගයකි. පර්යේෂකයින් කණ්ඩායම මෙය විකෘති kit RNA නිසා සිදුවන්නක් බව සොයා ගති. වෛරස වල ප්රවේනික ගබඩාව RNA වුවද සත්ව ලෝකය තුල ක්ෂීරපායින්ගේ මෙය ඉතා කලාතුරකින් සිදුවන්නකි.
ජානයක ක්රියාකාරී ඒකකය
දර්ශීය ඉයුකැරියෝටාවෙකුගේ ප්රෝටීන් වලින් වටවු ජානයක පින්තූරයක් DNA අණුවක කුමන කොටසක් M-RNA අණු බවට පත්කෙරෙන්නේ ද යන්න අනුග්රාහක හා තහවුරු කිරීමේ පොදස් මගින් තීරණය කරනු ලබයි. මුල් M-RNA අණුව පසුව පණිවිඩ RNA වලට කැඩෙන අතර ඒවා ප්රෝටීන බවට පත් වෙයි.
සෑම ජානයකම නියාමක පෙදෙස් ඇති අතර ඊට අමතරව පැහැදිලිවම ප්රෝටීන හා RNA වලට ෙක්ත සපයන පෙදෙස ඇත. සෑම ජානයකම ඇති නියාමක පෙදෙස් අනුග්රහක පෙදෙස් නම් වන අතර ජානයක් පිටපත් වී ප්රකාශ වීමට ආසන්න වූ විට පිටපත් වී ප්රකාශ අනුග්රහක පෙදෙස් වල එම ස්ථානයේ පොදු සමාන අනුපිළිවෙලක් ඇති ඒකමතික අනුපිලිවෙලවල් තිබුණද සමහර ජාන වලට ශක්තිමත් අනුග්රහක පෙදෙස ඇති අතර (පිටපත් යන්ත්රය සමග හොඳින් බැදෙන), සමහර ඒවා දුර්වලව බැදෙන දුර්වල අනුග්රාහක කොටස් හා බැදේ. පිටපත් කරන යාන්ත්රණය මෙම දුර්වල අනුග්රාහක කොටස් සමග බැෙඳන්නේ කලාතුරකින් නිසා ශක්තිමත් අනුග්රහකයින්ට වඩා මෙම දුර්වල අනුග්රාහකයින් පිටපත් කිරීමේ යෙදෙන්නේ සුළු වශයෙනි. අනෙකුත් නියාමක පෙදෙසක් වු තහවුරු කිරීමේ පෙදෙස් මෙම දුර්වල පෙදෙස් වෙනුවට යොදා ගනී. බොහෝ නියාමක පෙදෙස් එනම් පිටපත් කිරීම පටන් ගන්නා ස්ථානයන්හි 5 කොනට කලින් හෝ ඒ දෙසට පවතී. ඉයුකාරි යෝටාවන්ගේ අනුග්රාහක පෙදෙස් ප්රෝකාරීයෝටාවන්ට වඩා සංකිර්ණ හා අඳුනාගැනීමට අපහසුයි. බොහෝ ප්රෝකාරියොටාවන්ට වඩා සංකිර්ණ හා අඳුනාගැනීමට අපහසුයි.
බොහෝ ප්රෝකාරියෝටා ජාන ඔප්රන්ස් වලට සංවිධානගතවී ඇත. මේවා ජාන භාණ්ඩ වන අතර ඒවායේ නිෂ්පාදන වල ක්රියා එකිනෙකට නෑනම් කියන අතර ඒකකයක් ලෙස පිටපත් වේ. ප්රතිවිරුද්ධ ලෙස ඉයුකැරියෝටා ජාන වරෙකට එක බැගින් පිටත් වේ. එනමුත් ඉන්ට්රෝන නම් වු දිගු DNA කොටස් අඩංගු වන අතර ඒවා පිටපත් වන නමුත් කෙසේවත් ප්රෝටීන බවට පරිවර්තනය නොවේ. (පරිවර්තනයට පෙර ඒවා කැපී වෙන්වේ) මෙම කුඩා කැබලි වලට කැපීම / පැලීම ප්රෝකැරියාටාවන්ගේ ද ඉයුකාරි යොටාවන්ට වඩා බොහෝ අඩු සම්භාවිතයකින් සිදුවේ.
ප්රවේණි දේහ
ජීවියෙක්ගේ මුළු ජාන සංඛ්යාව ගෙනෝමය ලෙස හඳුන්වන අතර ඒවා ප්රවේණි දේහ එකක හෝ බොහෝමයක ගබඩා වී ඇත. යම්කිසි ජානයක ප්රවේණි දේහයක් ස්ථානගත වී ඇති ස්ථානය පථය නම් වේ. ප්රවේණි දේහ තනි ඉතා දිගු DNA හෙලික්සයක් වන අතර එහි ජාන දහස් ගණන් අඩංගුය. ප්රෝකැරියෝටා, බැක්ටීරියා ආකියාවන් දර්ශීය ලෙස ඔවුන්ගේ ජිනෝම තනි විශාල වලයාකාර ප්රවේණි දේහ තුළ තැන්පත් කරන අතර ඉතා කුඩා ජාන සංඛ්යාව අඩංගු වන අතර ජීවීන් අතර ලෙහෙස්ගේ මාරු කරගත හැක. උදා - ලෙස ප්රතිජීවකවලට ඔරොත්තු දෙන ජාන බැක්ටීරියා ප්ලාස්මිඩවල අඩංගු අතර තිරස් ජාන ජාන මාරුව මඟින් වෙන වෙනත් විශේෂ හරහාත් සෛල අතර මේවා මාරු වේ. සමහර සරල ඉයුකැරියෝටාවන් ද මෙලෙස ඉතා කුඩා ජාන සංඛ්යාවක් සහිතව ප්ලාස්මිඩ තබා ගත්ත ද බොහෝ ඉයුකැරියෝට ජාන ගබඩා වී ඇත්තේ දිගු ප්රවේණි දේහ බහු සංඛ්යාවක් තුළ හිස්ටොන් මඟින් බැඳ තබා ගන්නා සංකීර්ණයක් ලෙසට න්යෂ්ටියේ තැන්පත්වය. හිස්ටෝනවල DNA ගබඩාවන ආකාරයත්, හිස්ටෝනවල ඇති රසායනික වෙනස්කම් ද ජානයක් ප්රකාශ වීමට DNA වල කුමන පෙදෙසක් අදාල ද යන්න පාලනය කරන නියාමක ක්රියාවලීන් වේ. ඉයුකැරියෝටා ප්රවේණි දේහවල අන්ත නැවත නැවත යෙදෙන එකම අනුපිළිවෙලවල් සහිත ටෙලොමියර් නම් වූ කොටස්වලින් ආවරණය වී ඇත. මෙම ටෙලෝමියර් ජාන සඳහා කේත නොකරන අතර DNA බෙදීමේ දී කේත පෙදෙස හා නියාමක පෙදෙස් බිදී යාම වැළකීමට යෙදේ. සෛල බෙදීම සඳහා සුදානම් වීමේ දී කෙරෙන DNA බෙදෙන සෑම වාරයක් ගණනේම මෙම ටෙලෝමියරවල දි අඩුවේ. මෙම සංසිද්ධිය සෛල මහළු වීම සඳහා පිළිතුරක් ද සපයයි. එනම් මෙය සෛල ජාන බෙදීමට ඇති හැකියාවක් කෙමෙන් අඩු වී ගොස් ජීවියාගේ වයස්ගත වීම සිදු කරයි, යන්නය.
ප්රෝකැරියෝටාවන්ගේ ප්රවේණි දේහ තුළ ජාන පමණක්ම අඩංගු වන අතර ඉයුකැරියෝටාවන් ගේ ජනක් හිස් DNA නම් වූ කිසිම කෘත්යයක් ඉටු නොකරන කොටස ද අඩංගුය. ඒක සෛලික ඉයුකැරියෝටාවන්ගේ එම DNA කොටස් කුඩා සංඛ්යාවක් ඇති අතර සංකීර්ණ බහු සෛලිකයන්ගේ (මානවයන් ඇතුළුව) හඳුනාගත් කෘත්යයක් නැති DNA වලින් බොහෝමයක් මෙසේ වේ. නමුත් දැන් පෙනෙන අයුරින් මානව ජිනෝමයේ ප්රෝටීන කේත සඳහා ඇති ජාන 2%ක් පමණ වුව 80% පමණ වූ භෂ්ම ප්රකාශ වේ.
Gene expression
ප්රවේණික සංඥා
ජානයක් ප්රෝටීන බවට පරිවර්තනය කරන නීති කාණ්ඩයක් ලෙස ප්රවේනික කේත/ සංඥා හැඳින්විය හැක. සෑම ජානයක්ම DNA වල අඩංගු වන විශේෂිත නියුක්ලියෝටයිඩ අනුපිළිවෙලක් අඩංගුය. ජානයක් සාර්ථක ලෙස ක්රියාකාරී ප්රෝටීනයක් බවට පත්වීමට ප්රවේණි ද්රව්යයේ අච්චු ගල් වන නියුක්ලියෝටයිඩ හා ප්රෝටීනයේ තැනුම් ඒකක වන ඇමයිනෝ අම්ල ද අතර අනුරූපතාවක් තිබිය යුතුය. කෝඩෝන නම් වූ නියුක්ලියොටයිඩ 36න් කාණ්ඩ එක එකක් විශේෂිත ඇමයිනෝ අම්ලයකට හෝ සංඥාවකට අනුරූප වන අතර මෙම කොඩාන 3 “නැවතීමේ කොඩෝන” වේ. නව ඇමීනෝ අම්ලයක් සඳහා විශේෂිත වීමට අමතරව ජානයක අවසානය පැමිණ ඇති බවට පිටපත් කිරීමේ යාන්ත්රණයට ඒත්තු ගන්වයි. කොඩෝන 64ක් පමණ ඇති අතර (එක් එක් ස්ථාන 3 නියුක්ලියෝටයිඩ වර්ග 4කින් එනිසා 43 තිබිය හැකි කොඩෝන ගණවේ) ප්රමිතිගත ඇමයිනෝ අම්ල 20ක් පමණ ඇත්තේ එම නිසා සංඥා අතිරික්ත අතර සංඥා විශාල ගණනක් එකම ඇමයිනෝ අම්ලයට සංඥා කේත සපයයි. ඇමයිනෝ අම්ල හා කොඩෝන අතර ඇති අනුරූපතාවය සෑම ජීව සංවිධානයකට සමස්ථයකි.
ජාන පිටපත් කිරීම
ජාන පිටපත් කිරීමේ ක්රියාවලියේ දී පණිවිඩ RNA නම් වූ ඒක රැහැනකින් යුත් RNA අණු නිෂ්පාදනය වේ. එහි අනුපිළිවෙල පිටපත් වන DNA අණුවේ රැහැනට අනුරූප වේ. මෙම DNA රැහැන් සංඥා රැහැන් නම් වේ. RNA නිෂ්පාදනය වන DNA රැහැන අච්චු රැහැන වේ. RNA පොලිමරේස් එන්සයිමයෙන් පිටපත්වීම සිදු කරන අතර එහිදී අච්චු රැහැන 31 සිට 51 දිශාවට කියවන අතර RNA රැහැන 51 - 31 දිශාවට නිෂ්පාදනය වේ. පිටපත් වීම ඇරඹීමට එම එන්සයිමය මඟින් ජානයේ අනුග්රාහක පෙදෙස හඳුනාගත යුතුයි. එම නිසා ජාන පාලනයේ ප්රධාන යාන්ත්රණයක් වන්නේ අනුග්රාහක පෙදෙස හඳුනාගැනීමෙන් වැලකීම සඳහා අනුග්රාහක පෙදෙස සිර කිරීම, හෝ තනි කිරීම මඟිනි. (මෙය පොලිමරේස්ව කායිකව වළකන repressor නම් වූ මැඩ පවත්වන අණුවක් තදින් බැඳී හෝ DNA අණුව සංවිධානය වීමෙනි) ප්රෝකැරියෝටාවන්ගේ පිටපත්කරණය සෛල ප්ලාස්මයේ සිදු කෙරේ. දිගු පිටපත්වලට, පිටපත්කරණය 51 අන්තය පිටපත් වෙද්දීම RNA වල 51 අන්තයෙන් ද සිදුවේ. ඉයුකැරිහෝටාවරුන් මෙය න්යෂ්ටිය නම් වූ සෛලයේ DNA තනිව එක්රැස් වූ ස්ථානයේ දී සිදු වේ. ප්රථම පිටපත වු RNA අණුව පොලිමරේස් මඟින් ද නිපදවෙන අතර එය පශ්චාත් පිටපත් නවීකරණය සෛල ප්ලාස්මයට නිදහස්වීමට පෙර සිදු වේ.
පරිවර්තනය සඳහා ඉයුකැරියෝටාවන්ටම විශේෂිත ලෙස පිටපත් වූ පෙදෙසේ ඇති ඉන්ට්රෝන පුරුද්දීම සිදුවේ. විකල්ප පිරිද්දීම් යාන්ත්රණයක් මඟින් වැඩුණු පිටපත්වල එකම ජානයෙන් වුව ද වෙන වෙනත් අනුපිළිවෙලවල් ඇති අතර වෙන වෙනත් ප්රෝටීනවලට සංඥා සපයයි. ඉයුකැරියෝටා සෛලවල සිදුවන වැදගත් නියාමනයකි.
DNA විභාජනය හා ප්රවේණිගත වීම
ජීවියෙකු ගේ , වැඩීම , වර්ධනය ,වර්ගයා බෝකිරීම සෛල විභාජනය මත රඳාපවතී . එනම් තනි සෛලයකින් එකහා සමාන දුහිතෘ සෛල 2ක් බිහිකිරීම් ක්රියාවලියයි.මෙහිදී මුල්ම විභාජනය මගින් ජිනෝමයේ සෑම ජානයකින්ම පිටපත් සෑදීම කල යුතුය.මේ සඳහා පොලිමරේස් නම් විශේෂිත එන්සයිම අවශ්යයි. මෙය විසින් DNA ද්විත්ව හෙලික්සයේ එක් රුහැනක් කියවන අතර (එය පිටපත් කරන රුහැන නම් වේ) නව සමාන රුහැනක් නිෂ්පාදනය කරයි. DNA ද්විත්ව හෙලික්සය එකට තබා ගන්නේ භෂ්ම ගැලපීම නිසා බැවින් එක් රැහැනක අනුපිළිවෙල එහි අනෙක් රැහැනක අනුපිලිවෙල එහි අනෙක් රුහැනේ අනුපිළිවෙල තීරණය කරයි. එම නිසා එන්සයිම මගින් එක් රුහැනක් කියවීම ප්රමාණවත්ය. මෙම DNA විභාජනය ඒකගුණ එනම් දුහිතෘ සෛලවල අඩංගු වන්නේ එක් මාතෘ DNA රුහැනක් හා අලුතින් සෑදුනු DNA රැහැනකි.
DNA විභාජනය සම්පූර්ණ වූ පසු සෛලය කායිකව ජිනෝමය පටලයකින් වට වූ කොපි 2ක් ලෙසට ජිභෝමය වෙන්කරයි. ප්රෝකැරියෝටාවන් වූ බැක්ටීරියා , ඒකියාවන් මෙය සිදුවන්නේ Binary fission සරල ක්රියාදාමයකිනි. එහිදී එක් එක් වලයාකාර ජිභෝමයන් සෛල පටලයන්ට සම්බන්ධ වී දුහිතෘ සෛල බවට වෙන් වේ. එහිදී සෛල පටලය ඇතුලට නැවී සෛල ප්ලාස්මය පටලයකින් වටවූ කොටස් දෙකට බෙදී සිදු කරයි. ඉයුකැරියෝවන්ට වඩා ද්විඛණ්ඩනය ඉතා වේගවත්ය. යුකැරියෝටා සෛල විභාජනය සංකීර්න අතර සෛල චක්රය නම් වූ ක්රියාවලියක් S අවධ්යේදී DNA විභාජනයටද ,M අවධ්යේ දී ප්රවේනි දේහ (දුහිතෘ) හා සෛලප්ලාස්මය බෙදේ. සිස්ට් වැනි ඒක සෛලික ඉයුකැරියොවන් බොහොමයක අංකුරනය සිදුවේ. එහිදී සෛල ප්ලිස්මය සමාන නොවූ දෘහිතෘ ජීවීන්ට බෙදේ.
ජාන අණුක ප්රවේණිය
ජාන ප්රවේණි ද්රව්ය පිටපත් වී පරම්පරාවෙන් පරම්පරාවට සම්ප්රේෂණය වීම අණුක ප්රවේණියේ පදනම වේ. තවද ජාන වල අණුක හා පැරණිකම අතර සම්බන්ධයයි. ජීවසංවිධානයක් ඹවුන්ගේ මාතෘ ජීවීන්ට සමාන ලක්ෂණ ලබන්නේ දුහිතෘ ජීවියාගේ සෛලවල මාතෘ ජාන වල කොපි ලැබීම නිසාය. අලිංගික ප්රජනනය සිදුකරන ජීවීන්ගේ දුහිතෘ ජීවියා මාතෘ ජීවියාගේ ප්රවේණික කොපියක් හෝ ක්ලෝනයක් වේ. ලිංගික ප්රජනනය සිදුකරන ජීවීන්ගේ විශේෂිත සෛල විභාජනයක් වන ඌනනය මගින් ජන්මානු හෙවත් ජන්මානු සෛල (ඒකගුණ) නිපදවන අතර ඒවායේ ජානයක එක් කොපියක් පමණක් අඩංගු ඒක ගුණ නම් වේ. ස්ත්රි නිපදවන ජන්මාණු ඩිම්බ ලෙසත් පුරුෂ නිපදවන ජන්මාණු ශුක්රාණුත් නම් වේ. ජන්මාණු දෙකක් එක් වී සංසේචිත බිත්තරයක් සාදයි. එය ද්විගුණ වන එක් සෛලයකි. මවගෙන් හා පියාගෙන් එක බැගින් ලැබුණු කොපි 2ක් අඩංගුය. ඌනනයේ දී ජාන ප්රතිසංවිධානය / මංසල සෑදීම සමහර විට සිදු වේ මංසල තැනීමේ දී එක් ක්රොමැටිඩය නමින් හඳුන් වන එක් ප්රවේනි දේහයක එක් භාගයක ඇති DNA කොටසක් කැඩී අනෙක් ක්රොමැටිඩයේ ඇති DNA කොටසක් සමඟ හුවමාරු වේ. ක්රොමැටිඩ වල ඇති ඇලිල සමාන නම් කිසි වෙනසක් සිදු නොවේ. ක්රොමැටිඩ වර්ණදේහාංශ නම් වේ. මෙන්ඩල්ගේ මූලධර්මයට අනුව පීතෘ/මාතෘ ජාන වලින් ජණ්මාණු වලට යැවෙන්නේ කුමන කොපියයෙන්න අහඹුව සිදු වේ.
ජීවියකුගේ කුමන ඇලීලය යම් කිසි ලක්ෂණයක් උරුම කරයිද යන වග හා මොන ඇලීලය එය වෙනත් ලක්ෂයක් උරුම කරන්නේද යන්න අතර සම්බන්ධයක් නැත. ඇත්ත වශයෙන්ම මෙය සතය වන්නේ එකම ප්රවේණි දේහයක් මත නොමැති (වෙන වෙනත් ප්රවේනි දේහ වල ඇති) ජාන වලටත් නැතිනම් එකම ප්රවේනි දේහය බොහෝ දුරින් ඇති ජාන වලටත් පමණි. ජාන දෙකක් එකම ප්රවේණි දේහයක ලඟින් පිහිටන තරමට ජන්මාණු වලට ඒදෙකට එකට සම්ප්රේෂෂණය වීමේ සම්භාවිතාව වැඩිය. ඉතා ලඟින් පිහිටන ජාන දෙකක් මංසල තනා වෙන්වීම ඇත්තවශයෙන්ම කවදාවත් සිදුනොවන තරම්ය. හේතුව මංසල සංධි එම ජාන අතරින් සිදු නොවන බැවිනි. මෙය ප්රවේණික බන්ධය නම්වේ.
ජාන ඉතිහාසය

ජාන පවතින බවට පළමු වරට මතයක් ඉදිරිපත් කරන ලද්දේ ග්රෙගර් මෙන්ඩල් (1822 – 1884) විසිනි. ඔහු 1860 වර්ෂවලදී මෑ ශාකවල ආවේණික පිළිබද අධ්යයනය කරමින් ජනකයින්ගේ ලක්ෂණ ජනිතයින් වෙත රැගෙන යාමේ සාධකයක් පිළිබද කල්පිතයක් ඉදිරිපත් කරන ලදී. ඔහුගේ එක් පරීක්ෂණයක් සදහා වසර 10 කටත් වැඩි කාලයක් ගත කෙරිණි. මෙන්ඩල් විසින් ජාන යන වචනය භාවිතා නොකළ නමුත් ආවේනික ලක්ෂණ යන පදය භාවිතයෙන් තම පරීක්ෂණ ප්රතිඵල පැහැදිලි කරන ලදී. තවද ස්වාධීන සංරචකය , ප්රමුඛ හා නිලීන ලක්ෂණ අතර විශිෂ්ටතාවය , විෂම යුග්මක හා සමයුග්මක විශිෂ්ටතාව , පසු කලෙක එනමින් හදුන්වන ලද රූපාණු දර්ශ හා ප්රවේණිදර්ශ අතර විශිෂ්ටතාව යන කල්පිත ද පළමු වරට ඉදිරිපත් කරන ලද්දේ ග්රෙගර් මෙන්ඩල් පළමු වරට ඉදිරිපත් කරන ලද්දේ ග්රෙගර් මෙන්ඩල් විසිනි. 1889 වසරේ දී හියුගෝ ඩිව්රිස් විසින් මෙන්ඩල්ගේ සංකල්පයට නමක් යොදන ලදී. එම කාල වකවානුවේදී මෙන්ඩල්ගේ පරීක්ෂණ පිළිබද බොහෝවිට ඩිව්රිස් නොදැන සිටියා විය හැක. කෙසේ වුවත් ඩිව්රිස් විසින් අන්තඃසෙලීය ස්වර්ජනනය නැමැති තම පොතෙහි “එක් ආවේණික ලක්ණයක් රැගත් කුඩාම කොටස” යන අර්ථයෙන් “සර්වාංශු” යන වචනය ප්රබන්ධ කර තිබිණි. ඊට දශක දෙකකට පසුව විල්හෙල්ම් ජොනැන්සන් විසින් එම වචනය කෙටි කොට ජානය ලෙසින් නම් කරන ලදී.
1900 වසරවල මුල් කාලයේ දී මෙන්ඩල්ගේ පරීක්ෂණ කෙරෙහි විද්යාඥයින්ගේ අවධානය නැවත යොමු විය. 1910 වසරේදී තොමස් හන්ට් මෝර්ගන් විසින් විශේෂ වර්ණදේහ තුළ ජාන පවතින බව පෙන්වා දෙන ලදී. වර්ණදේහවල යම් විශේෂිත ස්ථානයන්හි ජාන පවතින බව පසු කලෙක දී ඔහු විසින් පෙන්වා දෙන ලදී. මේ කරුණු උපකාරයෙන් මෝර්ගන් සහ ඔහුගේ ශිෂ්යයින් පළමුවරට Drosophila නම් පළතුරු මැස්සාගේ වර්ණදේහ සිතියම් කිරීම ආරම්භ කරන ලදී. 1928 දී තමා විසින් සිදු කරන ලද පරීක්ෂණ පදනම් කොට ගෙන ජානවලට හුවමාරු විය හැකි බව ෆෙඩ්රික් ග්රිෆික් විසින් පෙන්වා දෙන ලදී. ඔහුගේ පරීක්ෂණයේදී තාපය හමුවේ අඩපණ කරන ලද මාරාන්තික බැක්ටීරියාවක් මීයෙකු තුළට එන්නත් කිරීමේ දී බැක්ටීරියාවේ අඩංගු වූ ප්රවේනික තොරතුරු ඒ හා සමාන මාරාන්තික නොවන බැක්ටීරියා සම හුවමාරු වීමෙන් මීයා මරණයට පත්විය. මෙම පරීක්ෂණය ග්රිෆිත්ගේ පරීක්ෂය ලෙසින් හදුන්වනු ලැබේ. ජාන විකෘති මගින් පරිවෘත්තීය ක්රියාවලින්ගේ සමහර පියවරවලදී ගැටලු ඇතිවන බව 1941 වසරේදී ජෝර්ජ් වෙල්ස් බීඩ්ල් සහ එඩ්වඩ් ලව්රි ටාටම් විසින් පෙන්වා දෙන ලදී. ඒක විශේෂිත ප්රෝටීනයකට එක් විශේෂිත ජාන සංඥා කේතයක් ඇති බව මෙයින් පැහැදිලි වේ. 1944 වසරේ දී ඔස්වල්ඩ් අව්රි කොලින් මැක්ලියෝ සහ මැක්ලින් මැකාටි විසින් ජානයක තොරතුරු DNA තුළ අඩංගු වන බව පෙන්වා දෙන ලදී. 1953 වසරේ දී ජේම්ස් . ඩී. වොට්සන් සහ ෆ්රැන්සිස් ක්රික් විසින් DNA හි අණුක ආකෘතිය ඉදිරිපත් කරන ලදී.
ජානයේ සංඝටක
වෙන වෙනත් ජීවීන්ගේ ජාන සංඛ්යාව [8] විශේෂය ජාන ගණන ප්ලස්මා ජෙනිටැලියම් 500 ස්ට්රොප්ටොකොකස් නියුමෝනියාව 2,300 එෂ්චරිසියා කෝලයි 4,400 සැකරෝමයිසිස් සර්ව්සියේ 5,800 ඩ්රොසෆිලා මෙලනොගැස්ටර් 13,700 සේනොහැබ්ඩයිට්ස් එලිගන්ස් 19,000 හොමෝ සේපියන්ස් 20,500[7] සී අර්වින් 23,300 ඇබිඩොප්සිස් කැලියානා 25,500 මස් මස්කියුලස් 29,000 ඔරයිසා සැටීවා 50,000
පරිනාමික ලෙස සමීප වුව ද බොහෝ ජීවී සංවිධානවල ජීනෝමයේ දර්ශීය ජාන සංඛ්යාව හා විශාලත්වය පුළුල් ලෙස වෙනස් වේ. මානව ජීනෝම ව්යාපෘතිය ඇරඹීමට පෙර මීයන්, පළතුරු මැස්සන්, ආදී වූ සරල සතුන්ට වඩා ඉතා විශාල ජාන සංඛ්යාවක් මිනිස් විශේෂවලට ඇතැයි යන්න විශ්වාස කළ ද එය ඇරඹූ පසු මානවයන්ගේ ජාන ඝනත්වය බොහෝ අඩු බව පෙනුණි. ජීනෝමයක ඇති ජාන සංඛ්යාවෙන් ඇස්තමේන්තු සකස් කිරීම අමාරු කාර්යයකි. එනම් එය ඉයුකැරියෝටාවන්ගේ TATA නියාමක පෙදෙස්, ඒකමතික අනුග්රාහක අනුපිළිවෙලට සමාන අනුග්රාහක ප්රදේශ විවෘත කියවන රාමු ආදී වූ ජාන සෙවීමට යොදා ගන්නා ඇල්ගොරිදම් මත තීරණය වන නිසාවෙනි. ප්රෝකැරියෝටාවන්ට වඩා ඉයුකැරියෝටාවන්ගේ ජීනෝමයේ ජාන සොයා ගැනීම අසීරුය. හේතුව ඉන්ට්රෝන බොරු ජාන නම් වූ කේත නොකරන DNA අඩංගු නිසාය. පරිගණක ජාන සෙවීම් ක්රම කළින් භාවිතා වූ යම් කිසි ඇලිලයකට (සොයාගත හැකි) මං විවර කළ විශේෂිත විකෘති සෑදූ ස්ථානයක් සෙවීමට ගත් සිතියම් ගත කිරීම්වලට වඩා මෙය විශ්වාසදායකය. බොහෝ ඉයුකැරියෝටාවන්ගේ විශේෂවල ජීනෝමයේ DNA කොටස් විශාල ප්රමාණයකින් ජාන වෙන් වන අතර මෙම කොටස් බොහෝමයක් ජීවියාට කිසිම කාර්යභාර්යක් ඉටු නොකරන junk DNA වේ. මෙම DNA වල බොරු ජාන නම් වූ ආකාරය පිළිබඳ අධ්යයනයන් කර ඇත. ඒවා ජාන ලෙසට ප්රකාශ වන නමුත් අත්යවශ්ය පරිදි අනුග්රාහක හා පාලක අනුපිළිවෙලවල් නැති කේත රහිත DNA කොටස්ය. අතීතයේ පරිනාමක පෙළපත්වල සිදු වූ ජාන පිටපත් කිරීමේ ප්රතිඵලයක් ලෙසට ඇති වූ පෙදෙස් ලෙස උපකල්පනය කරයි. තවදුරටත් ජාන කේත රහිත පෙදෙස වූ ඉන්ට්රෝන වලින් කොටස වී ඇති අතර එම ඉන්ට්රෝන කේත සහිත අනුපිළිවෙලවල්වලට වඩා බොහෝ විශාල විය හැකි අතර මුල් mRNA වල පිටපත් කළ පසු සිදුවන වෙනස්කම් වලදී මේවා කැපී ඉවත් වේ.
ජානයේ පරිණාමය පිළිබඳ සංකල්පය
හැඩ ගැසීම හා ස්වභාවික වරණය නම් 1966 දී පළ කළ ඔහුගේ පොතට අනුව ජෝජ් විලියම් ජානය පරිනාමයේ මධ්යස්ථානය ලෙස ඔහු සඳහන් කරයි. ස්භාවික වරණය මඟින් සමහර ජානවල කැමැත්තක් දක්වන විට දී ඔහු මෙම මතය ඉදිරිපත් කළේය. අර්ථ කථනය : වෙන් කිරීම හා ප්රති සංවිධානය පිළිගත හැකි සම්භාවිතාවයකින් සිදුවේ. මෙයට අනුව අලිංගික ජිනෝමයන් පවා ජානයක් ලෙස සැලකිය හැක. පරම්පරා ගණනක් තුරා එයට හොඳ පිළිගත හැකි සදාකාලික බවක් ලැබී ඇත. වෙනස නම් : අණුක ජානය ඒකකයක් ලෙස පිටපත් වන අතර පරිනාමික ජානය ඒකකයක් ලෙස උරුම වේ. වොර්ඩ් ඩ්වොකින්ස් “ආත්මාර්ථකාමී ජානය, පැතිර ගිය රූපාකාරය” මඟින් ජානයක් යනු ජිව පද්ධතිවල බෙදෙන එකම දේ ජානය පමණි යන්න අදහස ආරක්ෂා කරයි. මෙයින් අදහස් කෙරෙන්නේ ජාන පමණක් එම ව්යුහය බොහෝ දුරට නොවෙනස්ව සම්ප්රේශණය වන අතර කොපි ලෙසට යම් කිසි සදාකාලික බවක් ද ලබයි. එම නිසා වරණයේ ඒකක ජානය විය යුතුයි. ඩ්වොකින්ස් ඔහුගේ ආත්මාර්ථකාමී ජාන නම් වූ කෘතියෙන් ජානය නැවත අර්ථ කථනයක් කරන අතර එය උරුම වීමේ ඒකකය ලෙස (සාමාන්යයෙන් පවතින ලෙසට ජානයක් නම් DNA යනු DNA කොටසක් ප්රෝටීනයක් බවට කේත කරන්නක් බවට පවතින) හැඳින්වීමට උත්සාහ කරයි. River out of Eden නම් කෘතියෙන් ඔහු තවදුරටත් ඔහු ජානයේ මධ්ය - වරණය නැවත විග්රහ කරන අතර ජිවියෙක් තාවකාලික දේහ හෝ නොනැසීමට ඇති යන්ත්ර ලෙසට ජානවලට සේවය කරයි. භූ විද්යාත්මකව වෙන් කිරීම් වල දී මෙම ජාන ගංඟාව අන්තර් අභි ජනනයෙන් සරාවිහිතයෙක් බිහි නොකළ හැකි විශේෂ 2ක් තනමින් ශාඛා දෙකට බෙදේ.
ජාන ඉලක්ක කිරීම් සහ භාවිතා
ජාන ඉලක්ක කිරීම් ලෙස සාමාන්යයෙන් හඳුන්වන්නේ මීයන්ගේ ජාන වෙනස් කිරීමට හෝ බිද දැමීම යොදා ගන්නා තාක්ෂණික ක්රම හා කළල වර්ධනයේ දී ජාන කෙසේ ක්රියා කරයි ද යන්න මානව රෝගී තත්ව, වයසට යෑම හා අකර්මන්ය අධ්යයනය කිරීමට මීයන්ගේ ආකෘති සැපයීම හා යොදන ශිල්ප ක්රමයි. අකර්මන්ය මීයෙක් / knockout mice ලෙස හඳුන්වන්නේ එම මී ආකෘතියේ ජාන 1ක් හෝ බහුතරයක් ක්රියා විරහිත තත්වයකට පත් කළ මීයන්ටය. ඒක ගුණ ප්රති සංවිධානය (කලලයේ ක්රියා විරහිත තත්වයකට පත් කළ මීයන්ටය. ඒක ගුණ ප්රති සංවිධානය (කළලයේ වංශ සෛල Stem cell වල) මුලින්ම වාර්තාගත වූ අවධියේ සිට එය ජාන ඉලක්ක කිරීමට භාවිතා කෙරුණි. ජාන ඉලක්ක කිරීම ඉතා ශක්තිමත් තාක්ෂණයක් වන බවට ඔප්පු කර ඇති අතර ක්ෂීරපායී ජීනෝමය ඉතා නිවැරදි ලෙසට යොදාගෙන විකෘති සහිත මීයන් දස දහසක්වත් නිෂ්පාදනය කරන අතර විශේෂිත කාල වකවාණුවල විශේෂිත සෛලවල හෝ ඉන්ද්රියන්වල ක්රියාකාරී වන / කළ හැකි විකෘති ඇති කිරීම කළ හැක. මෙය වර්ධනය වන කාලයේ දීත් වැඩිහිටි සත්වයෙකුගේත් කළ හැක. ජාන ඉලක්ක කිරීමේ ශිල්ප ක්රමය ස්ථාන විකෘති (point mutations) සමාකාර මැනීම් විකෘති ඇලිල නිවරද්ය කිරීම්, ප්රවේණි දේහයේ DNA විශාල කොටසක ඇතුල් කිරීමේ හා මැකීමේ විකෘති ඇති කිරීම පටක විශේෂිත බිඳ දැමීම් අවකාශාත්මක හා කාලීන නියාමනයන්ට ද සම්බන්ධ කරන ආදී නවීකරණයන් දක්වා පුළුල්ව පැතිරී ඇත. අනුමානකරණ රූපාකාරයක් ඇති මී ආකෘතියක් තැනීමේ හැකියාවෙන් වර්ධනය, ප්රතිශක්තිකරණ විද්යාව, ස්නායු ජීව විද්යාව, පිළිකා විද්යාව, කායික විද්යාව, පරිවෘත්තීය හා මානව රෝගී තත්ව යන අංශවල අධ්යයනවල සෑම අවධියක දීම විශාල බලපෑමක් කළ හැකි බව අනුමාන කරයි. ඕනෑම සෛලයක් බවට විකරණය විය හැකි සෛල ලබා ගත හැකි ඕනෑම විශේෂයකට මෙය යෙදිය හැකි බවට මත දරයි. එම නිසා ගෘහස්ථ සතුන්, ශාකවල වැඩි දියුණු කිරීමවල ද යොදා ගත හැක.
ජාන ප්රකාශනය
සෑම ජිවියෙකුගේම ප්රෝටීනයක් එකම ප්රෝටීනයේ සංඥා සපයන ජානයෙන් වෙන් කිරීමට ප්රධාන පියවර 2හි 1. DNA සිට පණිවිඩ (mRNA) වලට එම DNA වල ඇති ජාන පිටපත් කරයි. 2. (mRNA) සිට ප්රෝටීනයට එය පරිවර්තනය විම. RNA වලට කේත සපයන ජාන ද පළමු පියවර හරහා යා යුතු නමුත් ප්රෝටීනවලට පරිවර්තනය නොවේ. මෙම ජීව විද්යාත්මකව ක්රියාකාරී RNA හෝ ප්රෝටීන අණුවක් නිපදවීමේ ක්රියාවලිය ජාන ප්රකාශ වීම නම් වේ. ඵලය ජාන නිෂ්පාදනයක් නම් වේ.
ප්රවේණිගත ද්රව්ය ජාන සංවිධාන
ජීවියෙක්ගේ මුළු ජාන සංඛ්යාව ගෙෙනා්මය ලෙස හඳුන්වන අතර ඒවා ප්රවේණි දේහ එකක හෝ බොහෝමයක ගබඩා වී ඇත. යම්කිසි ජානයක ප්රවේණි දේහයක් ස්ථානගත වී ඇති ස්ථානය පථය නම් වේ. ප්රවේණි දේහ තනි ඉතා දිගු DNA හෙලික්සයක් වන අතර එහි ජාන දහස් ගණන් අඩංගුය. ප්රෝකාරියෝටා, බැක්ටීරියා ඒකියාවන් දර්ශීය ලෙස ඔවුන්ගේ ජිනෝම තනි විහාල වලයාකාර ප්රවේණි දේහ තුළ තැන්පත් කරන අතර ඉතා කුඩා ජාන සංඛ්යාව අඩංගු වන අතර ජීවීන් අතර ලෙහෙස්ගේ මාරු කරගත හැක. උදා - ලෙස ප්රතිජීවකවලට ඔරොත්තු දෙන ජාන බැක්ටීරියා ප්ලාස්මිඩවල අඩංගු අතර තිරස් ජාන ජාන මාරුව මඟින් වෙන වෙනත් විශේෂ හරහාත් සෛල අතර මේවා මාරු වේ. සමහර සරල ඉයුකැරියෝටාවන් ද මෙලෙස ඉතා කුඩා ජාන සංඛ්යාවක් සහිතව ප්ලාස්මිඩ තබා ගත්ත ද බොහෝ ඉයුකැරියෝට ජාන ගබඩා වී ඇත්තේ දිගු ප්රවේණි දේහ බහු සංඛ්යාවක් තුළ හිස්ටොන් මඟින් බැඳ තබා ගන්නා සංකීර්ණයක් ලෙසට න්යෂ්ටියේ තැන්පත්වය. හිස්ටෝනවල DNA ගබඩාවන ආකාරයත්, හිස්ටෝනවල ඇති රසායනික වෙනස්කම් ද ජානයක් ප්රකාශ වීමට DNA වල කුමන පෙදෙසක් අදාල ද යන්න පාලනය කරන නියාමක ක්රියාවලීන් වේ. ඉයුකැරියෝටා ප්රවේණි දේහවල අන්ත නැවත නැවත යෙදෙන එකම අනුපිළිවෙලවල් සහිත ටෙලොමියර් නම් වූ කොටස්වලින් ආවරණය වී ඇත. මෙම ටෙලෝමියර් ජාන සඳහා කේත නොකරන අතර DNA බෙදීමේ දී කේත පෙදෙස හා නියාමක පෙදෙස් බිදී යාම වැළකීමට යෙදේ. සෛල බෙදීම සඳහා සුදානම් වීමේ දී කෙරෙන DNA බෙදෙන සෑම වාරයක් ගණනේම මෙම ටෙලෝමියරවල දි අඩුවේ. මෙම සංසිද්ධිය සෛල මහළු වීම සඳහා පිළිතුරක් ද සපයයි. එනම් මෙය සෛල ජාන බෙදීමට ඇති හැකියාවක් කෙමෙන් අඩු වී ගොස් ජීවියාගේ වයස්ගත වීම සිදු කරයි, යන්නය.
ප්රෝකැරියෝටාවන්ගේ ප්රවේණි දේහ තුළ ජාන පමණක්ම අඩංගු වන අතර ඉයුකැරියෝටාවන් ගේ ජනක් හිස් DNA නම් වූ කිසිම කෘත්යයක් ඉටු නොකරන කොටස ද අඩංගුය. ඒක සෛලික ඉයුකැරියෝටාවන්ගේ එම DNA කොටස් කුඩා සංඛ්යාවක් ඇති අතර සංකීර්ණ බහු සෛලිකයන්ගේ (මානවයන් ඇතුළුව) හඳුනාගත් කෘත්යයක් නැති DNA වලින් බොහෝමයක් මෙසේ වේ. නමුත් දැන් පෙනෙන අයුරින් මානව ජිනෝමයේ ප්රෝටීන කේත සඳහා ඇති ජාන 2%ක් පමණ වුව 80% පමණ වූ භෂ්ම ප්රකාශ වේ.