Baktériumok
| Evolúciós időszak: Archaikum vagy korábbi – jelenkor | ||
 Az Escherichia coli baktérium
25 000-szeres nagyításban | ||
| Rendszertani besorolás | ||
| ||
| Törzsek[1] | ||
Actinobacteria (sugárgombák) (magas G+C) Aquificae
Acidobacteria (acidobaktériumok) | ||
| Hivatkozások | ||
A Wikifajok tartalmaz Baktériumok témájú rendszertani információt. A Wikimédia Commons tartalmaz Baktériumok témájú kategóriát. |
A baktériumok (Bacteria) egysejtű, többnyire néhány mikrométeres sejtes mikroorganizmusok. Változatos megjelenésűek: sejtjeik gömb, pálcika, csavart alakúak lehetnek. A mikrobiológia egyik ága, a bakteriológia foglalkozik a baktériumok tudományos és élettani vizsgálatával.
A Föld minden élőhelyén megtalálhatóak a baktériumok: a vízben, a szárazföldön vagy a levegőben, még mélytengeri hőforrásokban és nukleáris hulladékban is.[2] Egy gramm talaj kb. 40 millió, egy milliliter felszíni víz egymillió baktériumsejtet tartalmaz. A Földön pedig összesen mintegy 5 kvintillió (5 × 1030) baktérium élhet.[3] A baktériumok alapvető szerepet töltenek be a bioszféra anyagforgalmában, mint például a légköri nitrogén megkötésében. Ennek ellenére a baktériumfajok nagy részét nem ismerjük: a baktériumtörzsek fele rendelkezik csak olyan fajokkal, amelyek laboratóriumi körülmények között tenyészthetők.[4]
Valamivel több baktérium van az emberi testben, mint emberi sejt. A legtöbb baktérium a bőr felszínén és az emésztőrendszerben található.[5] A baktériumok nagy része ártalmatlan vagy hasznos, de akad néhány fertőző megbetegedést kiváltó patogén (kórokozó) baktérium is, mint például a kolera, szifilisz, lépfene, lepra vagy a pestis kórokozója. Gyakori és súlyos bakteriális megbetegedés a tuberkulózis (TBC), amely évente kétmillió embert öl meg nagyrészt Afrikában, a Szaharától délre eső területeken. A fejlett országokban antibiotikumokat használnak a fertőzések leküzdésére. Ezek túlzásba vitt használata, különösképpen pedig a baktériumok széles körére ható antibiotikumok kiterjedt használata eredményeként egyre több antibiotikumellenálló típus fejlődött ki. Ennek egy speciális esete figyelhető meg a Clostridium difficile baktériumnál. Az antibiotikum-ellenállás elterjedéséhez hozzájárult ezeknek a gyógyszereknek a helytelen használata, az orvosi előírás pontos betartásának elhanyagolása (lásd lejjebb).
Az iparban a szennyvíztisztításban, a tejtermékek gyártásában, az antibiotikumok és más szerves anyagok előállításában használnak baktériumokat.[6]
A baktériumok prokarióta szervezetek, tehát szemben az állatokkal és más eukariótákkal, nincs sejtmagjuk és más membránnal határolt sejtszervecskéjük. Ámbár hagyományosan baktériumnak neveznek minden prokariótát, a tudományos nevezéktan az utóbbi pár évben megváltozott, miután molekuláris biológiai módszerekkel a prokariótákat sikerült két alapvetően eltérő felépítésű és származású csoportra különíteni. Ez a két domén az Archeák és a Bacteria.[7]
A bakteriológia története

Az első baktériumokat Anton van Leeuwenhoek[8] holland természettudós pillantotta meg 1674-ben, egy saját maga által készített egylencsés, kétszázszoros nagyításra képes mikroszkópban. Megfigyeléseit a Királyi Társasághoz írt leveleiben publikálta.[9][10] Maga a baktérium elnevezés a görög βακτηριον szóból származik, melynek jelentése „kis pálca”;[11] a nevet Christian Gottfried Ehrenberg javaslatára 1828-ban vezették be.
A 19. század második felében Louis Pasteur bizonyította a mikrobák szerepét az erjedéssel, rothadással és fertőzésekkel kapcsolatban. Pasteur nyomán Joseph Lister angol sebész 1865-ben felismerte, hogy a sebfertőzés okozói is baktériumok és orvosi műszereit karbolsavval sterilizálta. Ugyanebben az évben halt meg Semmelweis Ignác magyar szülész-nőgyógyász, aki 1847-ben állati eredetű „bomlott szerves anyag” nyílt sérülésekre való átvitelével magyarázta, és klórmeszes kézfertőtlenítéssel akadályozta meg a gyermekágyi láz (Streptococcus sebfertőzés) kialakulását.
Robert Koch 19. századi német orvosbiológus nevéhez fűződik számos baktériumfaj azonosítása. Különböző laboratóriumi technikái (például a lemeztenyészet) segítségével elkülönítette és azonosította a tuberkulózis, lépfene és kolera kórokozóját. A tuberkulózissal végzett kutatásaiért Koch 1905-ben Nobel-díjat kapott.[12] A Koch-féle posztulátumok – a betegségek mikrobiális eredetére vonatkozó követelmények – ma is használatban vannak.[13]
Habár már a 19. században ismert volt, hogy számos betegséget baktériumok okoznak, sokáig nem sikerült hatásos antibakteriális kezelést kidolgozni.[14] 1910-ben Paul Ehrlich fejlesztette ki az első antibiotikumot. A szifilisz kórokozóját, a Treponema pallidum nevű spirochaetát szelektíven festő anyagban cserélt ki komponenseket oly módon, hogy az új keverék a patogént szelektíven elpusztította.[15] Ehrlich szintén Nobel-díjat kapott az immunológia területén végzett munkájáért, és élen járt a különböző baktériumok kimutatására és azonosítására használt festési eljárások kidolgozásában. Az ő munkái képezték alapját a Gram-festésnek és a Ziehl–Neelsen-festésnek is.[16]
A baktériumok tanulmányozásában jelentős lépés volt Carl Woese azon felismerése 1977-ben, hogy az Archaeák a baktériumoktól eltérő evolúciós vonalat képviselnek.[17] Ez az új filogenetikus osztályozás a 16S riboszomális RNS szekvenálásán alapult, és a prokariótákat két evolúciós doménre osztotta, kialakítva így a 3 doménes rendszert.[18]
Eredet és evolúció
A mai baktériumok ősei egysejtű mikroorganizmusok, a Föld első életformái voltak, melyek 4 milliárd évvel ezelőtt éltek. Mintegy 3 milliárd éve az összes élőlény mikroszkopikus méretű volt, a baktériumok és Archaea domén ősi képviselői voltak az élet domináns formái.[19][20] Habár léteznek bakteriális kövületek, mint például a Sztromatolitok, a jellegzetes morfológiai jegyek hiánya nem teszi lehetővé, hogy a bakteriális evolúciót, vagy egy bizonyos baktériumfaj eredetét rajtuk keresztül lehessen tanulmányozni. A genetika azonban lehetővé teszi a bakteriális törzsfejlődés rekonstruálását, és ezek a kutatások arra utalnak, hogy a baktériumok az Archaea vonaltól elválva kezdtek el külön úton fejlődni.[21] A baktériumok és az Archaea utolsó közös ősei valószínűleg azok a termofil szervezetek lehettek, melyek 2,5–3,2 milliárd évvel ezelőtt éltek.[22][23]
A második nagy evolúciós szétválásban, az archeák és az eukarióták szétválásában is szerepet játszottak a baktériumok. Az eukarióták akkor jelentek meg, amikor ősi baktériumok endoszimbiózisra léptek az eukarióta sejtek őseivel, melyek maguk is feltehetően az Archea csoport tagjai voltak.[24] Ennek során az ősi forma bekebelezett egy alfa-proteobaktériumot (melyből később a mitokondrium lett) és egy cianobaktériumszerű organizmust (melyből később a színtest lett).[25][26] Ezt az ún. endoszimbionta-elméletet Lynn Margulis (1938–) amerikai kutató 1967-ben publikálta először.[27] Margulis szerint a bekebelezett kisebb prokarióta sejtek tovább éltek a sejten belül, és az együttélés sikeres sejtkapcsolatnak bizonyult. Az elmélet bizonyítéka lehet az, hogy a mitokondrium és a színtest bakteriális méretű; saját örökítőanyaggal rendelkeznek, ami a prokariótákhoz hasonlóan kör alakú DNS; saját enzimatikus apparátussal rendelkeznek és osztódásuk a sejt osztódásától független.
Morfológia

A baktériumok alakja és mérete nagyon változatos képet mutat. A baktériumsejtek az eukarióta sejteknél kb. 10-szer kisebbek, leggyakrabban 0,5–5 mikrométer a hosszúságuk. Azonban akad néhány faj, mint például a Thiomargarita namibiensis és a Epulopiscium fishelsoni, melyek akár a fél milliméteres nagyságot is elérik, és szabad szemmel is láthatóak.[28] A legkisebb baktériumok a Mycoplasma nemzetségbe tartozó fajok; mindössze 0,3 mikrométeres méretük megegyezik a legnagyobb vírusok méretével.[29]
A legtöbb baktériumfaj gömb vagy pálcika alakú. A gömb alakúak másik neve coccus a görög kókkos szó után, mely magot jelent. A pálcika alakúak másik neve bacilus, a latin baculus, pálca szóból származtatva. Tipikus képviselőjük a kólibacilus (Escherichia coli). Néhány pálcika alakú baktérium hajlott vessző alakú (más néven komma vagy vibrio alak), mint a koleravibrio (Vibrio cholerae). A spirillumok merev csavar alakú baktériumok. A dugóhúzó alakú, hosszú és nagyon vékony spirochaeták sejtfala nem merev, ezért mozgás közben elhajolnak. Kevés tetraéder vagy kocka alakú fajt is ismernek.[30] A baktériumok alakját a bakteriális sejtfal és a citoszkeleton (sejtváz) határozza meg. Az alak alapvetően befolyásolja, hogy a baktérium hogyan tud táplálékot szerezni, letapadni, folyadékban úszni, vagy támadói elől elmenekülni.[31][32] A sejtfallal eredetileg rendelkező, de azt elveszített, L-forma baktériumegyedek ezért a kiindulási baktérium alakjától függetlenül gömb vagy szferoid alakúak.
Számos baktériumfaj egyetlen sejtként éli le életét, mások jellegzetes mintázatot alkotva társulnak és csoportokat vagy telepeket képeznek egymással: a Neisseria fajok párokat (diploidokat) képeznek, a Streptococcusok láncot alkotnak, a Staphylococcusok szőlőfürtszerűen csoportosulnak. A baktériumok fonalszerűen megnyúlhatnak, mint például az Actinobacteria (sugárgombák). A fonál alakú baktériumokat gyakran tok veszi körül, mely számos egyedülálló sejtet is tartalmaz. Bizonyos fajok, mint a Nocardia nemzetség, összetett elágazó fonalakat formáz, mely megjelenésre hasonlít a gombák micéliumára.[33]

A baktériumok gyakran tapadnak különféle felületekhez és egybefüggő bevonatot, biohártyát (biofilm) vagy baktériumszőnyeget alkotnak. A bevonat vastagsága néhány mikrométertől a fél méterig terjedhet, és benne több baktériumfaj, valamint a Protista és az Archeák csoport képviselői is előfordulhatnak. A bevonatban élő baktériumok sejtjei és a sejten kívüli komponensek bonyolult módon rendeződnek el, másodlagos struktúrákat, például mikrokolóniákat hoznak létre, melyeken keresztül csatornák rendszere biztosítja, hogy a tápanyagok megfelelő módon jussanak el az egyes sejtekhez.[34][35] Természetes körülmények között, mint például a földben és a növények felületén a baktériumok többsége bevonatot alkotva található meg.[36] A biohártya fontos a krónikus bakteriális fertőzéseknél, vagy a beültetett orvosi eszközöknél fellépő fertőzéseknél, mert a baktériumokat megvédi, és így sokkal nehezebben pusztíthatók el, mint az egyedi sejtek.[37]
Néha még összetettebb morfológiai változások is lehetségesek. Aminosavhiány esetén a myxobaktérium (Myxobacteria) fajok sejtjei egymás felé vándorolnak, összetapadnak és akár 500 mikrométer hosszú, fajra jellemző alakú és színű termőtestet formáznak, melyekben közel 100 000 baktériumsejt található.[38] A termőtestben a baktériumok már külön feladatokat is végeznek a többsejtű szerveződés egyik egyszerű típusaként. Például kb. minden tizedik sejt a termőtest felszínére vándorol, és egy speciális állapotú sejtté, ún. myxospórává alakul. A myxospórák a kiszáradásnak és a káros környezeti feltételeknek jobban ellenállnak, mint a normális sejtek (kitartó képlet).[39]
Felépítés

Sejten belüli struktúrák
A bakteriális sejtet lipidmembrán, más néven sejtmembrán burkolja, mely egyrészt határolja a sejttartalmat, másrészt akadályt képez, és a tápanyagokat, fehérjéket és a citoplazma egyéb életfontosságú alkotórészeit a sejten belül tartja. A sejtmembrán szoros kapcsolatban áll a sejtet kívülről határoló sejtfallal. A foszfolipidekből és fehérjékből álló kettős hártya szerepe sokrétű: a DNS a mezoszómához tapad a membránon; a légzési enzimek is a membrán lemezes betüremkedéseiben helyezkednek el, illetve a bioszintetikus, metabolikus reakciók egy része is a hártya mentén folyik.
Mivel prokarióta szervezetek, nincsenek membránnal borított sejtszervecskék (sejtorganellumok) a citoplazmában, és így kevés sejten belüli struktúrát tartalmaznak. Mindegyikükből hiányzik a sejtmag, a mitokondrium, a színtest és az eukarióta sejtekben megtalálható többi sejtszervecske, mint például a Golgi-készülék vagy az endoplazmatikus retikulum.[40]
A baktériumok nem rendelkeznek membránnal borított sejtmaggal, örökítőanyaguk, a DNS rendszerint egy darab körkörös kromoszóma. Ez a citoplazmában levő szabálytalan formájú képletben, az ún. nukleoidban található,[41] a hozzátapadt hisztonszerű fehérjékkel és az RNS-sel együtt. A 80%-os víztartalommal rendelkező sejtplazmában találhatók (mint minden élő organizmus esetében) a fehérjeszintézist végző riboszómák, de ezek felépítése egyrészt eltér az eukarióták és az archeák riboszómáinak felépítésétől,[42] másrészt számuk jóval nagyobb, mint az eukariótákban. A Planctomycetes rend tagjai kivételesek abból a szempontból, hogy esetükben a nukleoidot membrán veszi körbe, és rendelkeznek egyéb membránnal borított sejtstruktúrákkal is.[43]
A baktériumok egy része sejten belüli tápanyag-raktározó gömböket (granulumokat) képez, melyek glikogént,[44] polifoszfátot,[45] esetleg ként[46] tartalmaznak. Ezek a granulumok lehetővé teszik, hogy a baktériumok ezeket az anyagokat későbbi használatra elraktározzák. Bizonyos baktériumfajok, mint például a fotoszintetizáló cianobaktérium-fajok gázvezikulumokat képeznek a sejten belül, melyekkel a sejtjeik felhajtóerejét szabályozzák annak érdekében, hogy optimális fény- és tápanyagviszonyok közé kerüljenek.[47]
Sejten kívüli struktúrák
A sejtmembránon kívül helyezkedik el a bakteriális sejtfal, mely a baktériumot védi a környezeti hatásoktól és esetleg a gazdaszervezet immunrendszere ellen. A sejtfal emellett fontos szerepet játszik a sejt magas ozmózisnyomásának fenntartásában, ami akár a légköri nyomás tizenötszöröse is lehet. A sejtfal fő alkotórésze peptidoglikán, azaz olyan molekulák, amelyekben a peptidekhez poliszacharidláncok kapcsolódnak kovalens kötéssel.[48] A bakteriális peptidoglikán (más néven murein) térhálós szerkezetű: poliszacharidláncai D-aminosavakat tartalmazó peptidekkel van keresztülkötve.[49] A bakteriális sejtfal eltér a növények és a gombák sejtfalától, mivel azok cellulózból, illetve kitinből állnak.[50] A baktériumok sejtfala az archeák sejtfalától is különbözik, mivel azok sejtfala nem tartalmaz peptidoglikánokat. A sejtfal alapvető fontossággal bír a túlélés szempontjából: a penicillinszármazékok éppen azáltal teszik lehetővé a baktériumok elpusztítását, hogy gátolják a peptidoglikán szintézisét.[50]
Leegyszerűsítve két különböző típusú sejtfal található a baktériumokban, ezek alapján Gram-pozitív és Gram-negatív baktériumokra lehet felosztani a fajokat. A név a baktériumfajok osztályozására régóta használatos Gram-festés eredményére utal.[51]
A Gram-pozitív baktériumok sejtfala vastag, sok peptidoglikán- és lipoteichnoinsav-réteget tartalmaz. A Gram-negatív baktériumok ezzel szemben viszonylag vékony sejtfallal rendelkeznek, mely csak néhány réteg peptidoglikánból áll, melyet lipopoliszacharidokat és lipoproteineket tartalmazó második lipidmembrán burkol. A legtöbb baktérium a Gram-negatív csoportba tartozik, csak a Firmicutes és Actinobacteria törzs tagjainak van Gram-pozitív sejtfala.[52] A felépítésbeli különbségek eltérő érzékenységet eredményeznek az antibiotikumokkal szemben, például a vankomicin csak Gram-pozitív baktériumokat tud elpusztítani, és Gram-negatív patogénekkel, mint például a Haemophilus influenzae vagy a Pseudomonas aeruginosa fajokkal szemben hatástalan.[53]
Számos baktérium esetében egy merev szerkezetű fehérjemolekulákból álló S-réteg borítja a sejtet.[54] Ez a réteg kémiai és fizikai védelmet biztosít a sejtfelszínnek, és egyben a makromolekulák diffúzióját akadályozza. Az S-rétegnek más, még kevéssé ismert funkciói is vannak. Ismeretes például, hogy a Campylobacter fertőzőképességéhez hozzájárul, és a Bacillus stearothermophilus esetében felszíni enzimeket is tartalmaz.[55]
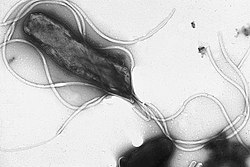
Az ostorok kb. 20 nanométer átmérőjű, és akár 20 mikrométer hosszúságú merev fehérjeképződmények, melyek az aktív helyváltoztatást szolgálják. A mozgáshoz szükséges energiát az elektrokémiai gradienst követve a sejtmembránon áthaladó ionok szolgáltatják.[56]
A csillók 2–10 nanométer átmérőjű és legfeljebb néhány mikrométer hosszú fehérjefonalak. A sejtfelszínt beborító csillók finom szőrzetre emlékeztetnek az elektronmikroszkópban. Mai ismereteink alapján a szilárd felületekhez vagy más sejtekhez történő tapadásban játszanak szerepet, és egyes patogén baktériumok fertőzőképességét is meghatározzák.[57]
A pilusok az ostoroknál némileg nagyobb sejtfüggelékek, melyeken keresztül az összetapadt baktériumsejtek genetikai anyagot cserélnek egymással (konjugáció, l. később).[58]
A baktériumok egy részét körülvevő tokok vagy nyálkaburkok szerkezetileg erősen eltérőek: megtalálható közöttük a sejten kívüli strukturálatlan polimertől kezdve a szigorúan strukturált tokig vagy glikokalix burokig minden. Ezek a struktúrák megvédhetik a sejteket más sejtek, például makrofágok által történő bekebelezéstől.[59] Antigénként szerepet játszhatnak abban, hogy az immunrendszer rajtuk keresztül felismeri a betolakodókat, de segítik a különböző felületekhez történő tapadást és a biohártyák képzését is.[60]
Ezeknek a sejten kívüli struktúráknak az összeállítása a bakteriális kiválasztórendszerektől függ. Ezek a rendszerek juttatják ki a fehérjéket a citoplazmából a periplazmába vagy a sejt környezetébe. Számos ilyen rendszer ismert, és mivel a patogének fertőzőképességének szempontjából meghatározóak, intenzíven kutatják ezeket.[61]
Endospórák

A Gram-pozitív baktériumok bizonyos nemzetségei, mint például a Bacillusok, Clostridiumok, például Clostridium difficile, Sporohalobacterek, Anaerobacterek vagy a Heliobacteriumok sejtjei nyugvó állapotú képletekké, ún. endospórákká alakulhatnak.[62] Legtöbbször csak egyetlen endospóra képződik a baktériumsejtben, amely az eredeti sejt pusztulásával kerül a szabadba, így célja nem a szaporodás, hanem a kedvezőtlen körülmények átvészelése (kitartó képlet). Az Anaerobacter fajok képesek akár 7 endospórát képezni egyetlen sejtben (kitartó és szaporító képlet).[63] Az endospórák közepén található a citoplazma a DNS-sel és a riboszómákkal, ezeket veszi körbe egy külső réteg (kéreg), melyet egy át nem eresztő merev burok zár körbe.
Az endospóráknak nincs anyagcseréje. Szélsőséges fizikai és kémiai körülményeket képesek átvészelni, például erős UV- vagy gamma-sugárzást, oldószereket, fertőtlenítőszereket, hőséget, nyomást és kiszáradást.[64] Sőt évmilliókig életképesek maradhatnak a nyugvó állapotban.[65][66] Az endospóráknak köszönhetően a baktérium még az űrben található vákuumot és sugárzást is túlélheti.[67] Az endospórákat képező baktériumok között kórokozók is akadnak: például a lépfene elkapható a Bacillus anthracis endospóráinak belélegzésével, vagy a mély sebbe jutott Clostridium tetani endospóra tetanuszt okoz.[68]
Anyagcsere
A magasabb rendű organizmusokkal szemben a baktériumok anyagcseréje nagyon változatos képet mutat.[69] Hagyományosan az anyagcsere jellegzetességei alapján határozták meg a rendszertanukat, de ez az osztályozás gyakran eltér a modern genetikai osztályozástól.[70] A bakteriális anyagcsere durva felosztásának alapját az adja, hogy az adott baktérium a növekedéshez milyen szén- és energiaforrást használ, valamint az energiatermelő folyamatok során mely anyagok és vegyületek adják az elektront (elektrondonor) és mely anyagok és vegyületek kapják a végén az elektront (elektronakceptor).[71]
Szénforrás szempontjából a baktériumok lehetnek heterotrófok, azaz a környezetükben található szerves szénvegyületeket használják szén- és energiaforrásként, vagy autotrófok, azaz szénforrásként a környezet szén-dioxidját használják. Az autotróf baktériumok tipikus képviselői a fotoszintetizáló cianobaktériumok, zöld kénbaktériumok és részben a bíborbaktériumok, de autotróf sok kemolitotróf faj is, mint például a nitrifikáló és a kénoxidáló baktériumok.[72]
Energiaforrás szempontjából a baktériumok vagy fotoszintetizálók, azaz fotoszintézis útján a fényből nyerik az energiát, vagy kemoszintetizálók, azaz kémiai vegyületekből nyerik az energiát. A kemoszintetizálókat tovább szokás bontani kemolitotrófokra (a légzéshez szervetlen elektrondonort használnak) és kemoorganotrófokra (a légzéshez szerves elektrondonort használnak). Kemolitotróf baktériumok esetében a leggyakoribb energiaforrás a hidrogén, szén-monoxid, ammónia (ennek eredménye a nitrifikálás), esetleg vasion, vagy más redukált fémion, és számos kénvegyület. A legtöbb kemolitotróf szervezet autotróf, míg a kemoorganotróf szervezetek heterotrófok.
Elektrondonorok és -akceptorok tekintetében: a kémiai vegyületek energiaforrásként történő felhasználása során az oxidálódó anyagból az elektronok a végső elektronfelvevőnek kerülnek átadásra, redukciós folyamat során. Ebben a reakcióban energia szabadul fel, mely az anyagcsere során felhasználható. Az aerob élőlények esetében az oxigén az elektronfelvevő. Anaerob élőlények esetében más szervetlen vegyület, például nitrát, szulfát, vagy szén-dioxid az elektronfelvevő, aminek eredménye az ökológiai szempontból is fontos denitrifikálás, kéntelenítés és acetogenezis[73][74]). Léteznek fakultatív anaerob baktériumok, melyek ha nem áll rendelkezésre végső elektronfelvevő, erjedéssel biztosítják életműködésüket. Ennek során cukrokból, vagy egyéb magas energiatartalmú vegyületekből állítanak elő az erjedés típusától függően tejsavat, etil-alkoholt, hidrogént, vajsavat vagy egyéb végtermékeket.
A környezetszennyezésre adott biológiai válaszban is fontosak ezek a folyamatok, például szulfátredukáló baktériumok termelik a környezetben található különösen mérgező higanyvegyületek (metil-, és dimetil-higany) nagy részét.[75] Az aerob fotoszintetizáló és a kemolitotróf szervezetek esetében oxigén az elektronfelvevő, de anaerob körülmények között az oxigén helyett szervetlen vegyületeket használnak.
Különleges eset a metanotróf baktériumok esete, amikor a metángáz szolgáltatja az elektronokat és egyben szénforrás is.[76]
A fotoszintézis során megkötött szén-dioxid mellett néhány baktérium a légköri nitrogént köti meg a nitrogenáz enzimmel (ilyenek például a talajban élő nitrifikáló baktériumok). A nitrogénkötő képesség csaknem mindegyik fent felsorolt anyagcseretípussal párosulhat.[77]
Szaporodás
A többsejtű szervezetektől eltérően a baktériumsejtek méretének növekedése és osztódása szorosan összefügg egymással. A baktériumok egy bizonyos méretig növekednek, majd kettéosztódnak.[78] A folyamat ivartalan szaporodás, amit a baktériumok esetében hasadásnak szokás nevezni, megkülönböztetve a valódi sejtmaggal és kromoszómákkal rendelkező eukarióta sejtek sejtosztódásától.[79] Optimális körülmények esetén a baktériumok rendkívül gyorsan növekednek és osztódnak, akár 9,8 perc alatt is megduplázódhat egy baktériumpopuláció.[80] A sejtosztódás során két azonos utódsejt keletkezik. Néhány ivartalanul szaporodó baktérium ennél bonyolultabb képleteket alakít ki a szaporodás során, ezek az újonnan létrejött utódsejtek eloszlását szabályozzák. Erre jó példa a myxobaktériumok termőteste, a Streptomyces fajok hifái vagy a bimbózás, mely során egy kitüremkedő rész letörik, és így jön létre az utódsejt.

Laboratóriumban a baktériumokat rendszerint szilárd vagy folyékony közegben tenyésztik. Tiszta tenyészetek izolálásához szilárd közeget, például agaragar-táptalajt, a szaporodás méréséhez vagy nagy mennyiségű sejt előállításához folyékony közeget használnak. A folyékony közeget folyamatosan keverik, hogy egyenletes sejtszuszpenziót kapjanak, amit könnyű tovább szaporítani és szállítani, viszont nehéz belőle egy-egy baktériumcsoportot elkülöníteni. A baktériumok azonosítása történhet szelektív (például bizonyos tápanyagok vagy antibiotikumok hozzáadásával vagy kihagyásával előállított) közeg felhasználásával.[81]
Nagy mennyiségű baktérium gyors és olcsó előállításához a legtöbb laboratóriumi technika bőségesen adagolja a tápanyagokat. Természetes körülmények között azonban a tápanyagok mennyisége véges, ami azt is jelenti, hogy a baktériumok nem tudnak korlátlanul szaporodni. A tápanyagok korlátossága különböző növekedési stratégiákhoz vezetett (r-K stratégia). Néhány organizmus rendkívül gyors szaporodásra képes, ha a tápanyagok rendelkezésre állnak (r-stratégia). Erre jó példa az algavirágzás jelensége, amely a nyári melegben oxigénszegénnyé vált, de tápanyagokban gazdag sekély tavakban katasztrofális méreteket is ölthet a cianobaktériumok (régi nevükön kékmoszatok) elszaporodásával.[82] Más baktériumok inkább utódaik túlélési esélyét növelik (K-stratégia). Például a Streptomyces fajok különféle antibiotikumokat termelnek, amivel más mikroorganizmusok növekedését gátolják.[83] A természetben sokan választják a közösségi életet (például biohártya), amely segíthet a táplálkozásban és védelmet is nyújthat,[36] de létfeltétel is lehet (például ilyen a szintrófia jelensége, amikor két mikroorganizmus kölcsönösen függ egymás anyagcseretermékétől).[84]
A baktériumpopulációk növekedése három fő szakaszra osztható. Amikor baktériumok kerülnek a megfelelő tápanyaggal ellátott környezetbe, a sejteknek először alkalmazkodniuk kell az új környezethez. A növekedés első szakasza a lappangó fázis, a lassú növekedés szakasza, mikor a sejtek felkészülnek és átállnak a gyors növekedésre a megfelelő enzimrendszerek, transzportfehérjék szintetizálásával.[85] A második növekedési szakasz a logaritmikus fázis, más néven exponenciális fázis. Ennek jellemzője a gyors, exponenciális növekedés. Az egyedszám időegység alatti növekedését mutatja a növekedési ráta, az egyedszám megduplázódását pedig a generációs idő. Ebben a fázisban a sejtek a tápanyagokat maximális sebességgel használják fel az anyagcseréjükben, a gyors reprodukció miatt a genetikai állomány megkettőződése folyamatosan zajlik. Még mielőtt az első kettőződés végbemenne, megkezdődik a következő. Ezért egy időben több replikációs villát is láthatunk a DNS-en. Ez egészen addig tart, míg a tápanyagok el nem kezdenek fogyni, korlátozva a szaporodást. Az utolsó fázis a stacioner vagy veszteglő fázis, melyet a tápanyaghiány okoz. A sejtek csökkentik az anyagcseréjüket, és lebontják a nem életfontosságú sejtfehérjéket. A stacioner fázis a gyors növekedés állapotából a stresszre adott válaszállapotba történő átmenet, melynek során megnövekedik a DNS-javítással, az antioxidáns-anyagcserével és a tápanyagszállítással összefüggő gének aktivitása.[86]
Genetika
A baktériumok többségének egyetlen, körkörös kromoszómája van. Méretét tekintve a Mycoplasma genitalium kórokozó 580 ezer bázispárral a legkisebb,[87] míg 12,2 millió bázispárral a talajlakó Sorangium cellulosum a legnagyobb[88] ismert bakteriális kromoszóma. A spirochaeták (például a Lyme-kór kórokozója, a Borrelia burgdorferi) ettől eltérően lineáris kromoszómával rendelkeznek.[89] A bakteriális kromoszóma hisztonok helyett hisztonszerű fehérjéket tartalmaz. A baktériumsejtben előfordulhatnak plazmidok is, olyan kis méretű, kör alakú öröklődő DNS-darabok, amelyek nem részei a kromoszómának. A plazmidok antibiotikum-rezisztenciáért, fertőzőképességért felelős géneket is hordozhatnak. A bakteriális DNS egy része víruseredetű. Számos bakteriális vírus, azaz bakteriofág ismeretes. Néhány egyszerűen megfertőzi és elpusztítja a baktériumokat, mások beépülnek a bakteriális kromoszómába. A bakteriofág tartalmazhat olyan géneket, melyek a gazda fenotípusát is befolyásolják. Például az Escherichia coli O157:H7 és a Clostridium botulinum evolúciója során bakteriofág toxingének változtatták át az eredetileg ártalmatlan baktériumokat halálos kórokozókká.[90][91]
A baktériumok ivartalanul szaporodnak, így utódaik genetikai állománya megegyezik. A baktériumok evolúciója a genetikai anyagban bekövetkezett rekombináció és mutáció révén előálló módosulások szelekciójával valósul meg. Mutáció a DNS hibás másolásakor, vagy mutagénekkel történő érintkezéskor következik be. A baktériumfajok, sőt az egy fajba tartozó törzsek mutációs rátája is nagyon eltérő lehet.[92] Mutációhoz vezethet a stressz is: ilyenkor bizonyos, a növekedést korlátozó folyamatokkal összefüggésben álló géneknek növekedik meg a mutációs rátája.[93]
Sok baktériumnál megfigyelték az örökítőanyag sejtek közötti átvitelét (horizontális géntranszfer). Ennek három fő módja van. A transzformáció során a baktérium képes a környezetében levő DNS-t felvenni. Az így felvett DNS gyakran nem kerül be a baktérium kromoszómájába, hanem plazmidként található meg a sejtben. Gének kerülhetnek be a baktériumba a transzdukció útján is, ekkor egy bakteriofág illeszt a bakteriális kromoszómába idegen DNS-t. A harmadik mód a konjugáció, amikor közvetlen sejtkapcsolat útján cserélődik ki DNS. A horizontális géntranszfer természetes körülmények között gyakori jelenség.[94] A génátvitelnek jelentős szerepe van az antibiotikum-rezisztencia szempontjából is, mivel lehetővé teszi a rezisztenciáért felelős gének gyors átadását akár különböző kórokozó fajok között is.[95]
Mozgás

A baktériumok ostorral, csúszással, rángatózó mozgással, vagy a felhajtóerő változtatásával képesek helyüket megváltoztatni.[96] A baktériumok között egyedülálló módon a spirochaetáknak ostorhoz hasonló képletei, ún. axiális filamentumai vannak, melyek nem a sejthártyában, hanem a sejthártya és a külső membrán közötti periplazmatikus térben találhatók. Jellegzetes spirálisan csavart testük van, mely mozgás közben meghajlik.[96]
Az ostorok száma és a sejt felszínén történő elrendeződése eltér a baktériumfajoknál. Vannak, melyeknek egyetlen ostora van (monotrich), léteznek fajok, melyeknél a sejt két végén van egy-egy ostor (amfitrich), esetleg a sejt egyik végén egy halomban van sok ostor (lofotrich), vagy a sejt mindkét végén több flagellum található (amfilofotrich), és ismertek fajok, melyeknél a sejt teljes felületét beborítják az ostorok (peritrich).

Az élőlények mozgását szolgáló valamennyi struktúra közül a baktériumok ostorának a szerkezetét és működését ismerjük a leginkább. Az ostor mintegy 20 fehérjéből épül fel, és körülbelül másik 30 fehérje játszik szerepet a szabályozásában és elkészülésében.[96] Az ostort az ostor tövében elhelyezkedő motor forgatja, amely a hajócsavarhoz hasonlóan hajtja előre a sejtet. Számos baktérium (például az E. coli) két különböző módon tud mozogni: előrehaladó mozgással (úszás) és bukfencezéssel. A bukfencezéssel tudnak új irányba állni, és térben mozogni.[97]
A mozgásra képes baktériumokat bizonyos ingerek vonzzák vagy taszítják, ezt a viselkedést taxis utótaggal jelölik: például kemotaxis, fototaxis vagy magnetotaxis.[98][99] A myxobaktériumoknál figyelhető meg az a jelenség, hogy az egyes baktériumsejtek együtt mozognak, miközben a sejtekből hullámok formálódnak, melyekből később az endospórákat tartalmazó termőtestek lesznek.[39] Ezek a baktériumok csak akkor mozognak, ha szilárd felszínen vannak, de például az E. coli akár folyékony közegben, akár szilárd felszínen is képes mozogni.
Néhány Listeria és Shigella faj a gazdasejten belül a gazdasejt citoszkeletonjának segítségével mozog (amit egyébként a sejt a sejtszervecskék mozgatására használ). A sejtjeik egyik oldalánál elősegítik az aktin polimerizációját, és a növekvő aktin filamentumok nyomják a másik irányba a baktériumsejteket a gazdasejten belül.[100]
Osztályozásuk és azonosításuk
| Linnaeus 1735 2 ország |
Haeckel 1866 3 ország[101] |
Chatton 1937 2 birodalom[102] |
Copeland 1956 4 ország[103] |
Whittaker 1969 5 ország[104] |
Woese et al. 1977 6 ország[105] |
Woese et al. 1990 3 domén[106] |
|---|---|---|---|---|---|---|
| - | Protista | Prokaryota | Monera | Monera | Eubacteria | Bacteria |
| Archaebacteria | Archaea | |||||
| Eukaryota | Protista | Protista | Protista | Eukarya | ||
| Vegetabilia | Plantae | Plantae | Fungi | Fungi | ||
| Plantae | Plantae | |||||
| Animalia | Animalia | Animalia | Animalia | Animalia |
Az osztályozás célja, hogy leírja az egyes baktériumfajok közötti eltéréseket az élőlények hasonlóságán alapuló csoportosítással és elnevezéssel. A baktériumok a sejtstruktúráik, anyagcseréjük vagy az olyan sejtalkotókban levő különbségek alapján osztályozhatóak, mint a DNS, zsírsavak, pigmentek, vagy az antigének.[81] Bár ezek alapján lehetővé vált a baktériumtörzsek azonosítása és osztályozása, nem volt világos, vajon a különbségek fajok közötti eltérések, vagy pedig egy fajon belüli eltérések. Ennek a bizonytalanságnak az az oka, hogy a legtöbb baktériumfajban nincsenek jól megfigyelhető jellegzetes képletek, valamint hogy az egymástól független fajok között is létezik a horizontális géntranszfer jelensége.[107] A horizontális géntranszfer miatt a közeli rokonságban álló baktériumoknak is egészen eltérő morfológiája vagy anyagcseréje lehet. Annak érdekében, hogy ez a bizonytalanság csökkenthető legyen, a modern bakteriológia egyre inkább a molekuláris rendszertanra támaszkodik, olyan technikákat felhasználva, mint a guanin-citozin arány meghatározása, vagy olyan gének DNS-szekvenciálása, melyeket nem érintett komolyan a horizontális géntranszfer (például rRNS-gének).[108]


A baktériumok azonosítása különösen az orvostudományban jut nagy szerephez, ahol a megfelelő kezelés a fertőzést okozó baktériumfaj ismeretétől függ. Emiatt a baktériumok azonosítására szolgáló technikák fejlődését döntően befolyásolta az emberi kórokozók azonosításának sürgető igénye.
A baktériumokat különböző festési eljárásokkal szembeni viselkedés alapján is szokták csoportosítani. Az egyik ilyen eljárás a Gram-festés, melyet 1884-ben Hans Christian Gram fejlesztett ki. Ez a módszer a baktériumokat a sejtfal strukturális sajátosságai alapján különíti el.[51] A festés során kristályibolya (vagy genciánaibolya)-festékkel festik meg a baktériumkészítményt, majd etanollal mosási próbát végeznek. Gram-pozitív baktériumok esetén a festék nem mosható ki a sejtből, míg a Gram-negatív baktériumoknál igen. A Gram-negatív baktériumok láthatóvá tétele érdekében további fukszinos festést alkalmaznak. A Gram-pozitív baktériumok lilának látszanak, míg a Gram-negatív fajok rózsaszínűek lesznek a festés után. A Gram-festést a morfológiával kombinálva a legtöbb baktérium besorolható 4 csoport egyikébe (Gram-pozitív coccusok, Gram-pozitív bacilusok, Gram-negatív coccusok, Gram-negatív bacilusok). Gram-pozitív baktériumok például a tüdőgyulladást okozó Streptococcus pneumoniae, Gram-negatív az emberi bélcsatornában élő Escherichia coli.
Bizonyos élőlények kimutatására a Gram-festés helyett más módszereket alkalmaznak, például a Mycobacterium tuberculosis vagy a Nocardia egyedeinek kimutatására elterjedt a Ziehl–Nielsen-féle saválló festés.[109] Ennél a módszernél forró fenolos fukszinnal festenek, ami ezekből a fajokból nem oldódik ki sav hatására (saválló), így a sejtek a háttérhez képest sötétebb színt vesznek fel. A Corynebacterium dyphtheriae foszfáttartalmú volutinszemcséit, az ún. Babeş–Ernst-szemcséket lehet láthatóvá tenni a Neisser-festéssel.
Számos baktériumot annak alapján azonosítanak, hogy milyen speciális közegen tenyészthetőek. Ezeknek a technikáknak az az alapja, hogy bizonyos baktériumfajok szaporodását a közeg elősegíti, míg más fajok szaporodását gátolja. Ezek a technikák gyakran bizonyos mintákra specifikusak, például a köpetmintát úgy kezelik, hogy a tüdőgyulladást okozó kórokozót azonosítsák, míg a székletmintákat olyan szelektív közegeken tenyésztik, hogy ki tudják mutatni a hasmenés kórokozóját, egyúttal megakadályozzák a nem kórokozó baktériumok szaporodását. A normális körülmények között steril mintákat (vér, vizelet, gerincvelői folyadék) olyan közegen tenyésztik, mely lehetővé teszi az összes baktérium szaporodását.[81][110]
Ezek mellett egyéb, például szerológiai technikákat is bevetnek azonosítás céljából.
Mint a baktériumok osztályozásánál, az azonosításnál is növekvő szerephez jutnak a molekuláris biológiai módszerek. Például gyorsan növekszik a tenyésztéshez képest gyors és pontos DNS alapú diagnosztikai módszer, a polimeráz-láncreakció népszerűsége.[111] Ezek a módszerek lehetővé teszik azoknak az élőlényeknek a felismerését és azonosítását is, amelyek ugyan anyagcseréjüket tekintve aktívak, azonban nem osztódnak, és nem tenyészthetőek kultúrában.[112]
Kapcsolat más élőlényekkel
Nyilvánvaló egyszerűségük dacára a baktériumok bonyolult módon képesek együttműködni más élőlényekkel. Ezeket a szimbionta kapcsolatokat feloszthatjuk parazita, mutualista és kommenzalista típusúakra. A baktériumok kis mérete miatt a kommenzalizmus közönséges, állatokon és növényeken ugyanolyan jól növekednek, mint bármilyen más felületen.
Mutualista együttműködés
A rizoszféra (a növényi gyökerek által befolyásolt talajrész) nitrifikáló baktériumai légköri nitrogént kötnek meg,[113] így könnyen felvehető nitrogént biztosítanak olyan növényeknek, amelyek maguktól nem képesek a molekuláris nitrogén megkötésére.
Számos más baktérium él szimbiózisban az emberrel és más élőlényekkel. A normál emberi bélflórában jelen levő körülbelül 1000-féle baktérium nagyban hozzájárul a bél immunrendszeréhez, folsav-, K-vitamin, és biotin-szintéziséhez, valamint a különben nem vagy nehezen emészthető szénhidrátok emésztéséhez.[114][115] Lactobacillus fajok segítik a tejfehérjék tejsavvá alakítását a bélben.[116] A bélflóra hasznos baktériumai gátolják a potenciálisan kórokozó baktériumok szaporodását (általában kompetitív kizárás révén). Ezeket a hasznos baktériumokat, mint probiotikus tápanyag-kiegészítőket forgalmazzák is.[117]
Kórokozók

Kórokozónak tekintjük, ha a baktériumok parazita együttműködést alakítanak ki más élőlénnyel. A fertőzés létrejöhet érintkezéssel, táplálékkal, levegő vagy víz útján.
A kórokozó baktériumoknak a vírusok mellett nagy szerepük van a betegségek, fertőzések kialakításában. Baktériumok állnak számos emberi megbetegedés mögött: különféle gyulladások, mint a mandulagyulladás, középfülgyulladás, szőrtüszőgyulladás, pattanás; gyomorrontást, hasmenést okozó fertőzések, mint a szalmonella, campylobacteriosis, vérhas, kolera; olyan ismert betegségek, mint a pestis, lepra, TBC (gümőkór), skarlát, diftéria, tífusz, tetanusz (vérmérgezés) és számos nemi betegség, mint a gonorrhea (tripper, kankó) vagy a vérbaj (szifilisz). Bizonyos betegségeknél sok évvel a betegség leírása után derül ki, hogy kórokozó áll a háttérben, erre példa, hogy sokáig nem volt egyértelmű, hogy a gyomorfekély hátterében a Helicobacter pylori baktérium áll. A növényi kórokozókra példa az almafélék tűzelhalásos megbetegedéséért felelős Erwinia amylovora baktérium.
Mindegyik kórokozó jellegzetes módon lép kapcsolatba a gazdaszervezettel. A Staphylococcus és Streptococcus például bőrfertőzést, tüdőgyulladást vagy akár szepszist okozhat.[118] Emellett ezek az élőlények a normális emberi flóra részei, a bőrön és az orrban is megtalálhatóak és semmiféle megbetegedést nem okoznak. Más baktériumok, például a csak más élőlények sejtjein belül növekedő és szaporodó Rickettsia fajok kivétel nélkül betegségeket okoznak (a Rickettsiák egyik faja okozza a tífuszt). A sejten belüli parazitákhoz tartoznak a Chlamydia fajok, melyek a tüdőgyulladásért vagy a húgyúti fertőzésekért felelősek, és talán a szívkoszorúér megbetegedésekben is részük van.[119] Végül léteznek az opportunista kórokozók (mint a Pseudomonas aeruginosa, Burkholderia cenocepacia, vagy a Mycobacterium avium), melyek csak akkor okoznak megbetegedést, ha a immunhiányos vagy cisztás fibrózisban szenvedő embereket fertőznek meg.[120][121]
A bakteriális fertőzések kezelhetőek antibiotikumokkal, melyeket baktericidnek nevezünk, ha elpusztítják a baktériumokat, vagy bakteriosztatikusnak hívunk, ha csak a szaporodásukat gátolja. Számos különböző típusú antibiotikum létezik, mindegyikük valamilyen módon gátol egy, a kórokozóban és a gazdaszervezetben eltérően működő folyamatot. Erre példa a chloramphenicol és a puromycin, melyek a bakteriális riboszómák működését gátolják, de nincsenek hatással az ettől eltérő eukarióta riboszómára.[122] Az antibiotikumokat az emberi megbetegedések kezelése mellett az intenzív állattenyésztés során is használják, annak érdekében, hogy serkentsék az állatok növekedését. Ezzel azonban hozzájárulhatnak a baktériumpopulációkban a gyors antibiotikum-ellenállás kialakulásához.[123]
A baktériumfertőzés elleni védekezés módjai

- Higiénia – Járvány idején kerülni kell a tömegeket, gyakran kell tisztálkodni, kezet mosni. Csak ivóvízből szabad inni, az élelmiszereket pedig lehetőleg frissen kell fogyasztani. Az orvosi műszereket használat előtt sterilizálni kell. Védőoltás, injekció beadása, sebészeti beavatkozás előtt a bőrt alkohollal kell fertőtleníteni.
- Jó kondíció – Kevésbé betegszünk meg, ha közérzetünk, testi, lelki állapotunk jó, ha eleget mozgunk, egészségesen táplálkozunk.
- Védőoltás – A baktériumos megbetegedések megelőzésének is eszköze lehet a védőoltás.
- Antibiotikumok – A már kialakult baktériumos megbetegedések ellenszere, baktériumölő, vagy szaporodását gátló anyag.
Figyelem: Ez egy kétélű fegyver; csak akkor segít, ha az orvosi előírást pontosan betartjuk, mert a gyógyszerkutatók a leghatásosabb gyógyszermennyiség, beszedési időszakok és összidőtartam összességét kísérletileg állapították meg úgy, hogy a kezelés a baktériumkoncentrációt annyira csökkentse, hogy a maradékkal egyéni immunrendszerünk könnyen elbánhasson. Ha a beteg, magát jobban érezve a gyógyszer bevételét elhanyagolja, vagy a kúrát nem fejezi be, akkor a baktériumok megmaradó hányada ismét felszaporulhat. Darwin természetes kiválasztódás elmélete szerint az az egyed marad meg, amely az adott körülményekhez gyorsabban alkalmazkodik. Itt a megmaradó baktériumok jobban ellenálltak a kezelésnek, vagyis akaratunk ellenére a kezelés elhanyagolásával egy antibiotikum-ellenálló csoportot fejlesztettünk ki.
Ügyeljünk tehát az orvos által előírt adagolás pontos betartására, hogy ne segítsük elő ellenálló baktériumok kifejlődését.
Jelentőségük az iparban és a technológiában
Az emberiség évezredek óta használja a baktériumokat (az élesztőkkel és penészgombákkal együtt) olyan alapvető élelmiszerek készítésére, mint a bor, sajtok, savanyúság, ecet, szójaszósz, savanyú káposzta vagy joghurt.[124][125]
Figyelemreméltó a baktériumoknak azon képessége, ahogy szerves anyagokat képesek lebontani. Emiatt az iparban hulladékfeldolgozásra, szennyvíztisztításra hasznosítják őket, de speciális szerves alapanyagú üveget is kristályosíttatnak baktériumokkal. A kőolajban levő szénhidrogéneket lebontó baktériumokat olajfoltok megszüntetésére használják.[126] Sikeresen alkalmazták ezt a módszert az 1989-es Exxon Valdez hajókatasztrófa olajszennyezésénél is, amikor műtrágyát szórtak ki a Prince William öbölben, hogy elősegítsék a természetesen is jelenlevő baktériumok szaporodását. Ipari szennyezőanyagok lebontására is használnak baktériumokat.[127] A vegyiparban gyógyszerek és mezőgazdasági termékek előállításában szintén baktériumok működnek közre.[128]
A biológiai védekezés során a növényvédőszerek helyett is használhatóak baktériumok. Leggyakrabban a Bacillus thuringiensis nevű Gram-pozitív talajlakó baktérium valamelyik alfaja jut szerephez, mivel ezek a Lepidoptera rendre (lepkék, molyok) specifikus „rovarirtók”.[129] Mivel az emberre, az élővilágra és a hasznos rovarokra kicsi vagy semmilyen káros hatással nincsenek, környezetbarát rovarirtónak tekinthetőek.[130][131]
A gyors növekedésük és a könnyű manipulálhatóságuk miatt a baktériumok a molekuláris biológia, a genetika és a biokémia „igáslovai”. A bakteriális DNS módosításával, és az ennek eredményeként megváltozó fenotípus tanulmányozásával a tudósok képesek a gének, enzimek funkcióját és az anyagcsere útjait meghatározni, és ezeket az eredményeket a magasabb rendű szervezetekre is alkalmazni.[132] A biotechnológiában a bakteriális anyagcsere és genetika megértése teszi lehetővé, hogy olyan módosított baktériumokat állítsanak elő, melyek terápiás célból képesek inzulin, növekedési faktorok vagy antitestek előállítására.[133][134]
Ajánlott irodalom
- Pesti, Miklós. Általános mikrobiológia. Budapest–Pécs: Dialóg Campus Kiadó (2001). ISBN 963-9123-71-4
- Gergely, Lajos. Orvosi mikrobiológia. Budapest: Semmelweis Kiadó (1999). ISBN 963-8154-93-4
- de Kruif, Paul. Bacillusvadászok. Budapest: Könyvbarátok Szövetsége (1931)
Jegyzetek
- ↑ Bacteria (eubacteria). Taxonomy Browser. NCBI. (Hozzáférés: 2008. szeptember 10.)
- ↑ Fredrickson J, Zachara J, Balkwill D, et al (2004). „Geomicrobiology of high-level nuclear waste-contaminated vadose sediments at the hanford site, washington state”. Appl Environ Microbiol 70 (7), 4230–41. o. [2008. szeptember 29-i dátummal az eredetiből archiválva]. PMID 15240306. (Hozzáférés: 2006. december 18.)
- ↑ Whitman W, Coleman D, Wiebe W (1998). „Prokaryotes: the unseen majority”. Proc Natl Acad Sci U S A 95 (12), 6578–83. o. [2008. március 5-i dátummal az eredetiből archiválva]. PMID 9618454. (Hozzáférés: 2006. december 18.)
- ↑ Rappé M, Giovannoni S. „The uncultured microbial majority”. Annu Rev Microbiol 57, 369–94. o. PMID 14527284.
- ↑ Sears C (2005). „A dynamic partnership: Celebrating our gut flora”. Anaerobe 11 (5), 247–51. o. PMID 16701579.
- ↑ Ishige T, Honda K, Shimizu S (2005). „Whole organism biocatalysis”. Curr Opin Chem Biol 9 (2), 174–80. o. PMID 15811802.
- ↑ Woese C, Kandler O, Wheelis M (1990). „Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya”. Proc Natl Acad Sci U S A 87 (12), 4576–9. o. PMID 2112744.
- ↑ Leeuwenhoek nevét illetően az Anton, Antony és Antonie van Leeuwenhoek írásforma is ismeretes
- ↑ Leeuwenhoek A (1753). „Part of a Letter from Mr Antony van Leeuwenhoek, concerning the Worms in Sheeps Livers, Gnats, and Animalcula in the Excrements of Frogs”. Philosophical Transactions (1683–1775) 22, 509–18. o. [2010. január 5-i dátummal az eredetiből archiválva]. (Hozzáférés: 2007. augusztus 19.)
- ↑ Leeuwenhoek A (1753). „Part of a Letter from Mr Antony van Leeuwenhoek, F. R. S. concerning Green Weeds Growing in Water, and Some Animalcula Found about Them”. Philosophical Transactions (1683–1775) 23, 1304–11. o. [2010. január 18-i dátummal az eredetiből archiválva]. (Hozzáférés: 2007. augusztus 19.)
- ↑ Etymology of the word “bacteria” Online Etymology dictionary.
- ↑ The Nobel Prize in Physiology or Medicine 1905 Nobelprize.org
- ↑ O'Brien S, Goedert J (1996). „HIV causes AIDS: Koch's postulates fulfilled”. Curr Opin Immunol 8 (5), 613–18. o. PMID 8902385.
- ↑ Thurston A (2000). „Of blood, inflammation and gunshot wounds: the history of the control of sepsis”. Aust N Z J Surg 70 (12), 855–61. o. PMID 11167573.
- ↑ Schwartz R (2004). „Paul Ehrlich's magic bullets”. N Engl J Med 350 (11), 1079–80. o. PMID 15014180.
- ↑ Biography of Paul Ehrlich Nobelprize.org
- ↑ Woese C, Fox G (1977). „Phylogenetic structure of the prokaryotic domain: the primary kingdoms”. Proc Natl Acad Sci U S A 74 (11), 5088–90. o. PMID 270744.
- ↑ Woese C, Kandler O, Wheelis M (1990). „Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya”. Proc Natl Acad Sci U S A 87 (12), 4576–79. o. PMID 2112744.
- ↑ Schopf J (1994). „Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic”. Proc Natl Acad Sci U S A 91 (15), 6735–42. o. PMID 8041691.
- ↑ DeLong E, Pace N (2001). „Environmental diversity of bacteria and archaea”. Syst Biol 50 (4), 470–78. o. PMID 12116647.
- ↑ Brown J, Doolittle W (1997). „Archaea and the prokaryote-to-eukaryote transition”. Microbiol Mol Biol Rev 61 (4), 456–502. o. PMID 9409149.
- ↑ Di Giulio M (2003). „The universal ancestor and the ancestor of bacteria were hyperthermophiles”. J Mol Evol 57 (6), 721–30. o. PMID 14745541.
- ↑ Battistuzzi F, Feijao A, Hedges S. „A genomic timescale of prokaryote evolution: insights into the origin of methanogenesis, phototrophy, and the colonization of land.”. BMC Evol Biol 4, 44. o. PMID 15535883.
- ↑ Poole A, Penny D (2007). „Evaluating hypotheses for the origin of eukaryotes”. Bioessays 29 (1), 74–84. o. PMID 17187354.
- ↑ Dyall S, Brown M, Johnson P (2004). „Ancient invasions: from endosymbionts to organelles”. Science 304 (5668), 253–7. o. PMID 15073369.
- ↑ McFadden G (1999). „Endosymbiosis and evolution of the plant cell”. Curr Opin Plant Biol 2 (6), 513–9. o. PMID 10607659.
- ↑ Lynn Sagan (1967). On the origin of mitosing cells. J. Theoretical Biology 14(3), 255–274. PMID 11541392
- ↑ Schulz H, Jorgensen B. „Big bacteria”. Annu Rev Microbiol 55, 105–37. o. PMID 11544351.
- ↑ Robertson J, Gomersall M, Gill P. (1975). „Mycoplasma hominis: growth, reproduction, and isolation of small viable cells”. J Bacteriol. 124 (2), 1007–18. o. PMID 1102522.
- ↑ Fritz I, Strömpl C, Abraham W (2004). „Phylogenetic relationships of the genera Stella, Labrys and Angulomicrobium within the 'Alphaproteobacteria' and description of Angulomicrobium amanitiforme sp. nov”. Int J Syst Evol Microbiol 54 (Pt 3), 651–7. o. [2008. október 10-i dátummal az eredetiből archiválva]. PMID 15143003. (Hozzáférés: 2006. december 27.)
- ↑ Cabeen M, Jacobs-Wagner C (2005). „Bacterial cell shape”. Nat Rev Microbiol 3 (8), 601–10. o. PMID 16012516.
- ↑ Young K (2006). „The selective value of bacterial shape”. Microbiol Mol Biol Rev 70 (3), 660–703. o. PMID 16959965.
- ↑ Douwes K, Schmalzbauer E, Linde H, Reisberger E, Fleischer K, Lehn N, Landthaler M, Vogt T (2003). „Branched filaments no fungus, ovoid bodies no bacteria: Two unusual cases of mycetoma”. J Am Acad Dermatol 49 (2 Suppl Case Reports), S170–3. o. PMID 12894113.
- ↑ Donlan R (2002). „Biofilms: microbial life on surfaces”. Emerg Infect Dis 8 (9), 881–90. o. PMID 12194761.
- ↑ Branda S, Vik S, Friedman L, Kolter R (2005). „Biofilms: the matrix revisited”. Trends Microbiol 13 (1), 20–26. o. PMID 15639628.
- ↑ a b Davey M, O'toole G (2000). „Microbial biofilms: from ecology to molecular genetics”. Microbiol Mol Biol Rev 64 (4), 847–67. o. PMID 11104821.
- ↑ Donlan RM, Costerton JW (2002). „Biofilms: survival mechanisms of clinically relevant microorganisms”. Clin Microbiol Rev 15 (2), 167–93. o. PMID 11932229.
- ↑ Shimkets L. „Intercellular signaling during fruiting-body development of Myxococcus xanthus.”. Annu Rev Microbiol 53, 525–49. o. PMID 10547700.
- ↑ a b Kaiser D. „Signaling in myxobacteria”. Annu Rev Microbiol 58, 75–98. o. PMID 15487930.
- ↑ Berg J., Tymoczko J. and Stryer L. (2002) Biochemistry. W. H. Freeman and Company ISBN 0-7167-4955-6
- ↑ Thanbichler M, Wang S, Shapiro L (2005). „The bacterial nucleoid: a highly organized and dynamic structure”. J Cell Biochem 96 (3), 506–21. o. PMID 15988757.
- ↑ Poehlsgaard J, Douthwaite S (2005). „The bacterial ribosome as a target for antibiotics”. Nat Rev Microbiol 3 (11), 870–81. o. PMID 16261170.
- ↑ Fuerst J (2005). „Intracellular compartmentation in planctomycetes”. Annu Rev Microbiol 59, 299–328. o. PMID 15910279.
- ↑ Yeo M, Chater K (2005). „The interplay of glycogen metabolism and differentiation provides an insight into the developmental biology of Streptomyces coelicolor”. Microbiology 151 (Pt 3), 855–61. o. PMID 15758231. (Hozzáférés: 2006. december 27.)
- ↑ Shiba T, Tsutsumi K, Ishige K, Noguchi T (2000). „Inorganic polyphosphate and polyphosphate kinase: their novel biological functions and applications”. Biochemistry (Mosc) 65 (3), 315–23. o. PMID 10739474.
- ↑ Brune DC. (1995). „Isolation and characterization of sulfur globule proteins from Chromatium vinosum and Thiocapsa roseopersicina”. Arch Microbiol 163 (6), 391–99. o. PMID 7575095.
- ↑ Walsby A (1994). „Gas vesicles”. Microbiol Rev 58 (1), 94–144. o. PMID 8177173.
- ↑ IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN) (1986). „Nomenclature of glycoproteins, glycopeptides and peptidoglycans”. Eur. J. Biochem. (159), 1–6. o.
- ↑ van Heijenoort J (2001). „Formation of the glycan chains in the synthesis of bacterial peptidoglycan”. Glycobiology 11 (3), 25R – 36R. o. PMID 11320055.
- ↑ a b Koch A (2003). „Bacterial wall as target for attack: past, present, and future research”. Clin Microbiol Rev 16 (4), 673–87. o. [2008. szeptember 28-i dátummal az eredetiből archiválva]. PMID 14557293. (Hozzáférés: 2006. december 27.)
- ↑ a b Gram, HC (1884). „Über die isolierte Färbung der Schizomyceten in Schnitt- und Trockenpräparaten”. Fortschr. Med. 2, 185–189. o.
- ↑ Hugenholtz P (2002). „Exploring prokaryotic diversity in the genomic era”. Genome Biol 3 (2), REVIEWS0003. o. PMID 11864374.
- ↑ Walsh F, Amyes S (2004). „Microbiology and drug resistance mechanisms of fully resistant pathogens.”. Curr Opin Microbiol 7 (5), 439–44. o. PMID 15451497.
- ↑ Engelhardt H, Peters J (1998). „Structural research on surface layers: a focus on stability, surface layer homology domains, and surface layer-cell wall interactions”. J Struct Biol 124 (2–3), 276–302. o. PMID 10049812.
- ↑ Beveridge T, Pouwels P, Sára M, Kotiranta A, Lounatmaa K, Kari K, Kerosuo E, Haapasalo M, Egelseer E, Schocher I, Sleytr U, Morelli L, Callegari M, Nomellini J, Bingle W, Smit J, Leibovitz E, Lemaire M, Miras I, Salamitou S, Béguin P, Ohayon H, Gounon P, Matuschek M, Koval S (1997). „Functions of S-layers”. FEMS Microbiol Rev 20 (1–2), 99–149. o. PMID 9276929.
- ↑ Kojima S, Blair D. „The bacterial flagellar motor: structure and function of a complex molecular machine”. Int Rev Cytol 233, 93–134. o. PMID 15037363.
- ↑ Beachey E (1981). „Bacterial adherence: adhesin-receptor interactions mediating the attachment of bacteria to mucosal surface”. J Infect Dis 143 (3), 325–45. o. PMID 7014727.
- ↑ Silverman P (1997). „Towards a structural biology of bacterial conjugation”. Mol Microbiol 23 (3), 423–9. o. PMID 9044277.
- ↑ Stokes R, Norris-Jones R, Brooks D, Beveridge T, Doxsee D, Thorson L (2004). „The glycan-rich outer layer of the cell wall of Mycobacterium tuberculosis acts as an antiphagocytic capsule limiting the association of the bacterium with macrophages”. Infect Immun 72 (10), 5676–86. o. [2008. szeptember 29-i dátummal az eredetiből archiválva]. PMID 15385466. (Hozzáférés: 2006. december 27.)
- ↑ Daffé M, Etienne G (1999). „The capsule of Mycobacterium tuberculosis and its implications for pathogenicity”. Tuber Lung Dis 79 (3), 153–69. o. PMID 10656114.
- ↑ Finlay B, Falkow S (1997). „Common themes in microbial pathogenicity revisited”. Microbiol Mol Biol Rev 61 (2), 136–69. o. PMID 9184008.
- ↑ Nicholson W, Munakata N, Horneck G, Melosh H, Setlow P (2000). „Resistance of Bacillus endospores to extreme terrestrial and extraterrestrial environments”. Microbiol Mol Biol Rev 64 (3), 548–72. o. PMID 10974126.
- ↑ Siunov A, Nikitin D, Suzina N, Dmitriev V, Kuzmin N, Duda V. „Phylogenetic status of Anaerobacter polyendosporus, an anaerobic, polysporogenic bacterium”. Int J Syst Bacteriol 49 Pt 3, 1119–24. o. PMID 10425769. (Hozzáférés: 2006. december 28.)
- ↑ Nicholson W, Fajardo-Cavazos P, Rebeil R, Slieman T, Riesenman P, Law J, Xue Y (2002). „Bacterial endospores and their significance in stress resistance”. Antonie Van Leeuwenhoek 81 (1–4), 27–32. o. PMID 12448702.
- ↑ Vreeland R, Rosenzweig W, Powers D (2000). „Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal”. Nature 407 (6806), 897–900. o. PMID 11057666.
- ↑ Cano R, Borucki M (1995). „Revival and identification of bacterial spores in 25- to 40-million-year-old Dominican amber”. Science 268 (5213), 1060–4. o. PMID 7538699.
- ↑ Nicholson W, Schuerger A, Setlow P (2005). „The solar UV environment and bacterial spore UV resistance: considerations for Earth-to-Mars transport by natural processes and human spaceflight”. Mutat Res 571 (1–2), 249–64. o. PMID 15748651.
- ↑ Hatheway C (1990). „Toxigenic clostridia”. Clin Microbiol Rev 3 (1), 66–98. o. PMID 2404569.
- ↑ Nealson K (1999). „Post-Viking microbiology: new approaches, new data, new insights”. Orig Life Evol Biosph 29 (1), 73–93. o. PMID 11536899.
- ↑ Xu J (2006). „Microbial ecology in the age of genomics and metagenomics: concepts, tools, and recent advances”. Mol Ecol 15 (7), 1713–31. o. PMID 16689892.
- ↑ Zillig W (1991). „Comparative biochemistry of Archaea and Bacteria”. Curr Opin Genet Dev 1 (4), 544–51. o. PMID 1822288.
- ↑ Hellingwerf K, Crielaard W, Hoff W, Matthijs H, Mur L, van Rotterdam B (1994). „Photobiology of bacteria”. Antonie Van Leeuwenhoek 65 (4), 331–47. o. PMID 7832590.
- ↑ Zumft W (1997). „Cell biology and molecular basis of denitrification”. Microbiol Mol Biol Rev 61 (4), 533–616. o. PMID 9409151.
- ↑ Drake H, Daniel S, Küsel K, Matthies C, Kuhner C, Braus-Stromeyer S (1997). „Acetogenic bacteria: what are the in situ consequences of their diverse metabolic versatilities?”. Biofactors 6 (1), 13–24. o. PMID 9233536.
- ↑ Morel, FMM, Kraepiel AML, Amyot M (1998). „The chemical cycle and bioaccumulation of mercury”. Annual Review of Ecological Systems 29, 543–566. o.
- ↑ Dalton H (2005). „The Leeuwenhoek Lecture 2000 the natural and unnatural history of methane-oxidizing bacteria”. Philos Trans R Soc Lond B Biol Sci 360 (1458), 1207–22. o. PMID 16147517. [halott link]
- ↑ Zehr J, Jenkins B, Short S, Steward G (2003). „Nitrogenase gene diversity and microbial community structure: a cross-system comparison”. Environ Microbiol 5 (7), 539–54. o. PMID 12823187.
- ↑ Koch A (2002). „Control of the bacterial cell cycle by cytoplasmic growth”. Crit Rev Microbiol 28 (1), 61–77. o. PMID 12003041.
- ↑ szerk.: Törő Imre: Az élet alapjai. Budapest: Gondolat Kiadó (1989). ISBN 9632821815
- ↑ Eagon R. „Pseudomonas natriegens, a marine bacterium with a generation time of less than 10 minutes”. J Bacteriol 83, 736–7. o. PMID 13888946.
- ↑ a b c Thomson R, Bertram H (2001). „Laboratory diagnosis of central nervous system infections”. Infect Dis Clin North Am 15 (4), 1047–71. o. PMID 11780267.
- ↑ Paerl H, Fulton R, Moisander P, Dyble J. „Harmful freshwater algal blooms, with an emphasis on cyanobacteria”. ScientificWorldJournal 1, 76–113. o. PMID 12805693.
- ↑ Challis G, Hopwood D. „Synergy and contingency as driving forces for the evolution of multiple secondary metabolite production by Streptomyces species”. Proc Natl Acad Sci U S A 100 Suppl 2, 14555–61. o. [2008. július 25-i dátummal az eredetiből archiválva]. PMID 12970466. (Hozzáférés: 2006. december 29.)
- ↑ Kooijman S, Auger P, Poggiale J, Kooi B (2003). „Quantitative steps in symbiogenesis and the evolution of homeostasis”. Biol Rev Camb Philos Soc 78 (3), 435–63. o. PMID 14558592.
- ↑ Prats C, López D, Giró A, Ferrer J, Valls J (2006). „Individual-based modelling of bacterial cultures to study the microscopic causes of the lag phase”. J Theor Biol 241 (4), 939–53. o. PMID 16524598.
- ↑ Hecker M, Völker U. „General stress response of Bacillus subtilis and other bacteria”. Adv Microb Physiol 44, 35–91. o. PMID 11407115.
- ↑ Fraser C, Gocayne J, White O, Adams M, Clayton R, Fleischmann R, Bult C, Kerlavage A, Sutton G, Kelley J, Fritchman R, Weidman J, Small K, Sandusky M, Fuhrmann J, Nguyen D, Utterback T, Saudek D, Phillips C, Merrick J, Tomb J, Dougherty B, Bott K, Hu P, Lucier T, Peterson S, Smith H, Hutchison C, Venter J (1995). „The minimal gene complement of Mycoplasma genitalium”. Science 270 (5235), 397–403. o. PMID 7569993.
- ↑ Pradella S, Hans A, Spröer C, Reichenbach H, Gerth K, Beyer S (2002). „Characterisation, genome size and genetic manipulation of the myxobacterium Sorangium cellulosum So ce56”. Arch Microbiol 178 (6), 484–92. o. PMID 12420170.
- ↑ Hinnebusch J, Tilly K (1993). „Linear plasmids and chromosomes in bacteria”. Mol Microbiol 10 (5), 917–22. o. PMID 7934868.
- ↑ Brüssow H, Canchaya C, Hardt W (2004). „Phages and the evolution of bacterial pathogens: from genomic rearrangements to lysogenic conversion”. Microbiol Mol Biol Rev 68 (3), 560–602. o. PMID 15353570.
- ↑ Perna N, Mayhew G, Pósfai G, Elliott S, Donnenberg M, Kaper J, Blattner F (1998). „Molecular evolution of a pathogenicity island from enterohemorrhagic Escherichia coli O157:H7”. Infect Immun 66 (8), 3810–7. o. PMID 9673266.
- ↑ Denamur E, Matic I (2006). „Evolution of mutation rates in bacteria”. Mol Microbiol 60 (4), 820–7. o. PMID 16677295.
- ↑ Wright B (2004). „Stress-directed adaptive mutations and evolution”. Mol Microbiol 52 (3), 643–50. o. PMID 15101972.
- ↑ Davison J (1999). „Genetic exchange between bacteria in the environment”. Plasmid 42 (2), 73–91. o. PMID 10489325.
- ↑ Hastings P, Rosenberg S, Slack A (2004). „Antibiotic-induced lateral transfer of antibiotic resistance”. Trends Microbiol 12 (9), 401–4. o. PMID 15337159.
- ↑ a b c Bardy S, Ng S, Jarrell K (2003). „Prokaryotic motility structures”. Microbiology 149 (Pt 2), 295–304. o. [2007. szeptember 14-i dátummal az eredetiből archiválva]. PMID 12624192. (Hozzáférés: 2007. július 1.)
- ↑ Wu M, Roberts J, Kim S, Koch D, DeLisa M (2006). „Collective bacterial dynamics revealed using a three-dimensional population-scale defocused particle tracking technique”. Appl Environ Microbiol 72 (7), 4987–94. o. [2008. szeptember 29-i dátummal az eredetiből archiválva]. PMID 16820497. (Hozzáférés: 2006. december 30.)
- ↑ Lux R, Shi W (2004). „Chemotaxis-guided movements in bacteria”. Crit Rev Oral Biol Med 15 (4), 207–20. o. PMID 15284186.
- ↑ Frankel R, Bazylinski D, Johnson M, Taylor B (1997). „Magneto-aerotaxis in marine coccoid bacteria”. Biophys J 73 (2), 994–1000. o. PMID 9251816.
- ↑ Goldberg MB (2001). „Actin-based motility of intracellular microbial pathogens”. Microbiol Mol Biol Rev 65 (4), 595–626. o. PMID 11729265.
- ↑ E. Haeckel. Generelle Morphologie der Organismen. Reimer, Berlin (1866)
- ↑ E. Chatton. Titres et travaux scientifiques. Sette, Sottano, Italy (1937)
- ↑ H. F. Copeland. The Classification of Lower Organisms. Palo Alto: Pacific Books (1956)
- ↑ R. H. Whittaker (1969). „New concepts of kingdoms of organisms”. Science 163, 150–160. o.
- ↑ C. R. Woese, W. E. Balch, L. J. Magrum, G. E. Fox and R. S. Wolfe (1977). „An ancient divergence among the bacteria”. Journal of Molecular Evolution 9, 305–311. o.
- ↑ Woese C, Kandler O, Wheelis M (1990). „Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya.”. Proc Natl Acad Sci U S A 87 (12), 4576–9. o. DOI:10.1073/pnas.87.12.4576. PMID 2112744.
- ↑ Boucher Y, Douady CJ, Papke RT, Walsh DA, Boudreau ME, Nesbo CL, Case RJ, Doolittle WF (2003). „Lateral gene transfer and the origins of prokaryotic groups.”. Annu Rev Genet 37, 283–328. o. PMID 14616063.
- ↑ Olsen G, Woese C, Overbeek R (1994). „The winds of (evolutionary) change: breathing new life into microbiology”. J Bacteriol 176 (1), 1–6. o. PMID 8282683.
- ↑ Woods G, Walker D (1996). „Detection of infection or infectious agents by use of cytologic and histologic stains”. Clin Microbiol Rev 9 (3), 382–404. o. PMID 8809467.
- ↑ Weinstein M (1994). „Clinical importance of blood cultures”. Clin Lab Med 14 (1), 9–16. o. PMID 8181237.
- ↑ Louie M, Louie L, Simor AE (2000). „The role of DNA amplification technology in the diagnosis of infectious diseases”. CMAJ 163 (3), 301–309. o. PMID 10951731.
- ↑ Oliver J. „The viable but nonculturable state in bacteria”. J Microbiol 43 Spec No, 93–100. o. [2007. szeptember 28-i dátummal az eredetiből archiválva]. PMID 15765062. (Hozzáférés: 2007. január 22.)
- ↑ Barea J, Pozo M, Azcón R, Azcón-Aguilar C (2005). „Microbial co-operation in the rhizosphere”. J Exp Bot 56 (417), 1761–78. o. PMID 15911555. (Hozzáférés: 2006. december 31.)
- ↑ O'Hara A, Shanahan F (2006). „The gut flora as a forgotten organ”. EMBO Rep 7 (7), 688–93. o. PMID 16819463.
- ↑ Zoetendal E, Vaughan E, de Vos W (2006). „A microbial world within us”. Mol Microbiol 59 (6), 1639–50. o. PMID 16553872.
- ↑ Gorbach S (1990). „Lactic acid bacteria and human health”. Ann Med 22 (1), 37–41. o. PMID 2109988.
- ↑ Salminen S, Gueimonde M, Isolauri E (2005). „Probiotics that modify disease risk”. J Nutr 135 (5), 1294–8. o. PMID 15867327.
- ↑ Fish D. „Optimal antimicrobial therapy for sepsis”. Am J Health Syst Pharm 59 Suppl 1, S13–9. o. PMID 11885408.
- ↑ Belland R, Ouellette S, Gieffers J, Byrne G (2004). „Chlamydia pneumoniae and atherosclerosis”. Cell Microbiol 6 (2), 117–27. o. PMID 14706098.
- ↑ Heise E. „Diseases associated with immunosuppression”. Environ Health Perspect 43, 9–19. o. PMID 7037390.
- ↑ Saiman, L. „Microbiology of early CF lung disease”. Paediatr Respir Rev. volume = 5 Suppl A, S367–369. o. PMID 14980298
- ↑ Yonath A, Bashan A (2004). „Ribosomal crystallography: initiation, peptide bond formation, and amino acid polymerization are hampered by antibiotics”. Annu Rev Microbiol 58, 233–51. o. PMID 15487937.
- ↑ Khachatourians G (1998). „Agricultural use of antibiotics and the evolution and transfer of antibiotic-resistant bacteria”. CMAJ 159 (9), 1129–36. o. PMID 9835883.
- ↑ Johnson M, Lucey J (2006). „Major technological advances and trends in cheese”. J Dairy Sci 89 (4), 1174–8. o. PMID 16537950.
- ↑ Hagedorn S, Kaphammer B. „Microbial biocatalysis in the generation of flavor and fragrance chemicals”. Annu Rev Microbiol 48, 773–800. o. PMID 7826026.
- ↑ Cohen Y (2002). „Bioremediation of oil by marine microbial mats”. Int Microbiol 5 (4), 189–93. o. PMID 12497184.
- ↑ Neves L, Miyamura T, Moraes D, Penna T, Converti A. „Biofiltration methods for the removal of phenolic residues”. Appl Biochem Biotechnol 129–132, 130–52. o. PMID 16915636.
- ↑ Liese A, Filho M (1999). „Production of fine chemicals using biocatalysis”. Curr Opin Biotechnol 10 (6), 595–603. o. PMID 10600695.
- ↑ Aronson A, Shai Y (2001). „Why Bacillus thuringiensis insecticidal toxins are so effective: unique features of their mode of action”. FEMS Microbiol Lett 195 (1), 1–8. o. PMID 11166987.
- ↑ Bozsik A (2006). „Susceptibility of adult Coccinella septempunctata (Coleoptera: Coccinellidae) to insecticides with different modes of action”. Pest Manag Sci 62 (7), 651–4. o. PMID 16649191.
- ↑ Chattopadhyay A, Bhatnagar N, Bhatnagar R (2004). „Bacterial insecticidal toxins”. Crit Rev Microbiol 30 (1), 33–54. o. PMID 15116762.
- ↑ Serres M, Gopal S, Nahum L, Liang P, Gaasterland T, Riley M (2001). „A functional update of the Escherichia coli K-12 genome”. Genome Biol 2 (9), RESEARCH0035. o. PMID 11574054.
- ↑ Walsh G (2005). „Therapeutic insulins and their large-scale manufacture”. Appl Microbiol Biotechnol 67 (2), 151 – 9. o. PMID 15580495.
- ↑ Graumann K, Premstaller A (2006). „Manufacturing of recombinant therapeutic proteins in microbial systems”. Biotechnol J 1 (2), 164 – 86. o. PMID 16892246.
Fordítás
- Ez a szócikk részben vagy egészben a Bacteria című angol Wikipédia-szócikk fordításán alapul. Az eredeti cikk szerkesztőit annak laptörténete sorolja fel. Ez a jelzés csupán a megfogalmazás eredetét és a szerzői jogokat jelzi, nem szolgál a cikkben szereplő információk forrásmegjelöléseként.
További információk
- Magyar Mikrobiológiai Társaság
- Képek baktériumokról (angol gyűjtemény)
- Baktérium definíció a Pallasból
- A baktériumok sejtfala
- Baktériumok - Kertészeti lexikon, tuja.hu