Wandelende takken
| Wandelende takken Fossiel voorkomen: Krijt[1] – heden | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|

| |||||||||||
| De Spaanse wandelstok (Pijnackeria hispanica), een exemplaar uit Frankrijk | |||||||||||
| Taxonomische indeling | |||||||||||
| |||||||||||
| Orde | |||||||||||
| Phasmida Leach, 1815 | |||||||||||

| |||||||||||
| Een variatie van Extatosoma tiaratum is nagenoeg onzichtbaar tussen de met korstmossen begroeide takken, afgebeeld is een onvolwassen vrouwtje. | |||||||||||
| Afbeeldingen op | |||||||||||
| Wandelende takken op | |||||||||||
| |||||||||||
Wandelende takken,[2] ook wel wandelende takken en bladeren genoemd[3] (Phasmida), zijn een relatief kleine orde van insecten. Wandelende takken komen wereldwijd voor in tropische en subtropische bossen, een klein aantal leeft in gematigde streken. Alle soorten leven uitsluitend van bladeren en zijn 's nachts actief, ze schuilen overdag tussen de planten. Het is een vrij jonge groep waarvan de vertegenwoordigers bekend zijn van fossielen uit het Krijt.
Wandelende takken kennen een breed scala aan lichaamsvormen en -kleuren. Er zijn verschillende soorten die zeer groot kunnen worden: sommige bereiken een totale lichaamslengte van meer dan 50 centimeter. Veel soorten hebben een glad lichaamsoppervlak maar andere bezitten stekels en bulten waardoor ze beter opgaan in de omgeving waarin ze leven. Veel soorten hebben een zeer dun en langwerpig lichaam. Een uitzondering vormen de wandelende bladeren, die juist een breed en bladachtig lichaam hebben. Wandelende takken vertonen een breed scala aan camouflage en lijken vaak sterk op al dan niet dorre takken of bladeren, zodat ze voor hun natuurlijke vijanden moeilijk te ontdekken zijn.
Wandelende takken vertonen vaak een passieve vorm van verdediging door zich stil te houden of zich te laten vallen bij verstoring. Er zijn echter ook soorten die zich effectief weten te verweren tegen vijanden. Vrouwtjes kunnen vaak levensvatbare eieren produceren zonder te paren met een mannetje. Hier komen in de regel alleen vrouwtjes uit. Bij sommige soorten zijn er zelfs helemaal geen mannetjes bekend. Er zijn meer dan 3000 vertegenwoordigers, die verdeeld worden in veertien families.
Naamgeving
De wetenschappelijke naam Phasmida is afgeleid van het geslacht Phasma, dat op zijn beurt weer een Latinisering is van het Griekse woord voor (spook)verschijning; φάσμα (phasma). In de Engelse taal worden de dieren aangeduid met verschillende namen, waarvan stick insects (takinsecten) en walking sticks (lopende takken) het bekendst zijn. In het Duits is de naam gespenstschrecken het meest gebruikt, dat vrij vertaald 'spooksprinkhanen' betekent. In andere talen is de naam een variatie op de wetenschappelijke naam Phasmida, zoals het Spaanse fásmidos en het Franse phasmes. Voor de wandelende takken werden in het verleden verschillende wetenschappelijke namen gebruikt zoals Cheleutoptera, Phasmatoptera, Phasmoptera en Phasmatodea, waarvan de laatste het bekendste is en veel in de literatuur wordt gebruikt.
De naam Phasmida voor de groep van wandelende takken werd voor het eerst geopperd door William Elford Leach in 1815. De naam Phasmatodea werd pas later voorgesteld door Georgij Georgiewitsch Jacobson en Valentin Lvovich Bianchi in 1902. Vaak wordt in de taxonomie de eerst beschreven naam aangehouden. De naam Phasmida is bovendien korter dan Phasmatodea en lijkt het meest op het Engelse woord voor de insecten; 'phasmids'.[4] Er is echter ook kritiek op de naam Phasmida omdat deze naam grammaticaal niet juist is.[5] Aangezien de naam van de orde is afgeleid van het genus Phasma zou de uitgang Phasmatida moeten zijn en niet Phasmida. Daarom gebruiken sommigen liever de oudst gebruikte naam met de juiste uitgang; Phasmatodea. Hierdoor worden beide namen in de literatuur door elkaar gebruikt.
Wandelende takken werden in 1758 door Linnaeus ingedeeld in het geslacht Gryllus, dat tegenwoordig een groep van krekels is, maar indertijd nog werd ingedeeld bij de kevers (Coleoptera).[4] Later deelde hij de wandelende takken samen met de bidsprinkhanen in bij het geslacht Mantis. Wandelende takken werden voor het eerst in 1825 als een aparte groep beschouwd door Pierre André Latreille, die ze tot de rechtvleugeligen rekende als de familie Phasmidae. De vroegste classificatie van de groep werd in 1908 voorgesteld door Karl Brunner-von Wattenwyl en Josef Redtenbacher. Albert Günther presenteerde in 1953 een herziene indeling van de wandelende takken, waarin de groep voor het eerst als een aparte orde werd gezien onder de naam Phasmatodea. Uit een onderzoek van Paul Flook en Charles Hugh Fraser Rowell in 1998 bleek door DNA-sequencing dat wandelende takken het nauwst verwant zijn aan de rechtvleugeligen.[6]
Verspreiding en habitat

Wandelende takken hebben een zeer groot verspreidingsgebied en komen vooral voor in warme klimaten. Ze ontbreken alleen in noordelijke delen van Europa, Azië en Noord-Amerika, op Antarctica en delen van Patagonië.
Wereldwijd
Wandelende takken komen wereldwijd van nature voor van ongeveer 45° noorderbreedte tot ongeveer 45° zuiderbreedte; verreweg de meeste soorten leven in de tropen. Vooral in delen van Zuidoost-Azië en Zuid-Amerika zijn veel soorten te vinden. Daarnaast komen ze voor in zuidelijk Noord-Amerika, Afrika, het Midden-Oosten, Midden-Azië, Oceanië en Australië.[7]
Europa
In Europa komen relatief weinig vertegenwoordigers voor, hier zijn ongeveer twintig soorten bekend.[8] Voorbeelden zijn soorten uit de geslachten Bacillus, Clonopsis en Leptynia. Clonopsis gallica komt voor tot in de Franse regio Bretagne. In het Middellandse Zeegebied is de soort Leptynia hispanica algemeen. Ook in het noordelijk gelegen Engeland komen drie soorten voor, maar deze zijn geïntroduceerd en allemaal afkomstig uit Nieuw-Zeeland.[8] In Cornwall, Devon en Dorset leeft de soort Acanthoxyla inermis, in Cornwall en Devon is Acanthoxyla geisovii te vinden en de wandelende tak Clitarchus hookeri is aangetroffen op de Scilly-eilanden. Al deze soorten zijn meegekomen met de import van Nieuw-Zeelandse planten. Ze leven dan ook exclusief van planten die voorkomen in dit land, zoals pohutukawa en manuka.[9]
België en Nederland
In Nederland en België komen van nature geen soorten voor. Wel worden regelmatig wandelende takken waargenomen die in de natuur zijn gedumpt, voornamelijk exemplaren van de Indische wandelende tak (Carausius morosus). Eieren van deze soort werden in 1897 door Franse missionarissen naar Nederland gestuurd, waar ze werden opgekweekt door pater Robert de Sinéty (1872–1931). Waarschijnlijk zijn alle populaties van deze soort in westelijk Europa afkomstig uit het nageslacht van deze dieren.
Begin 2020 werd door het EIS Kenniscentrum Insecten echter vastgesteld dat er een stabiele populatie van Clonopsis gallica in Nederland voorkomt. Het is hiermee de eerste vertegenwoordiger van de insectenorde die zich hier heeft gevestigd.[10] De locatie werd geheim gehouden om het wegvangen van de dieren te voorkomen.[11]
Habitat
De habitat van wandelende takken bestaat hoofdzakelijk uit tropische en subtropische bossen, zowel in laaglanden als in bergstreken, en tropische en subtropische droge scrublands en bossen. Daarnaast komen ze voor in tropische en subtropische vochtige scrublands, gematigde bossen en scrublands, en tropische en subtropische droge graslanden. Ook in door de mens aangepaste streken, zoals landelijke tuinen, parken, groenstroken en wegbermen kunnen de dieren worden aangetroffen.[12] In Afrika komen wandelende takken vooral voor in graslanden; deze soorten hebben een zeer lang en sprieterig lichaam waarmee ze niet opvallen tussen de grashalmen.
Wandelende takken leven in verschillende vegetatielagen, maar zowel deze lagen als de soorten die erin leven kennen een sterke overlap. In de kruidlaag op de bosbodem en het onderste deel van de struiklaag tot een hoogte van ongeveer vijf meter komen kleine tot middelgrote soorten voor, die zowel gevleugeld als ongevleugeld kunnen zijn. Voorbeelden zijn soorten uit de geslachten Dares, Pylaemenes, Haaniella, Acanthoclonia, Parectatosoma, Ramulus, Lonchodes en Pseudophasma. In struikachtige planten in het bovenste deel van de struiklaag en het onderste deel van de boomlaag tot een hoogte van ongeveer tien meter komen de grotere en veelal vliegende soorten voor, zoals die uit de geslachten Orthonecroscia, Calvisia, Necroscia, Diesbachia en Hermarchus. In tropische regenwouden komen bomen voor van vijftien tot meer dan vijftig meter hoogte. Deze laag wordt wel de emerging layer (Engels voor 'kruinlaag') genoemd. Hier leven veelal de kleinere vertegenwoordigers van de wandelende takken zoals de soorten uit de geslachten Necroscia, Nescicroa, Stratocles, Prisopus en Phyllium.[7]
Uiterlijke kenmerken

P = Prothorax
Mo = Mesothorax
Ma = Metathorax
A = Abdomen
An = Antenne
O = Oog
Co = Coxa
Tr = Trochanter
Fe = Femur
Ti = Tibia
Ta = Tarsus
Sp = Sporen
Ce = Cerci
Wandelende takken staan bekend als extreem goed gecamoufleerde insecten. Ze lijken op verschillende plantendelen, zoals takjes, bladeren, uitlopers, stukken schors of met mos begroeide takjes. Soorten die van grassen leven hebben vaak een zeer dun en langwerpig lichaam. Soorten in tropische regenwouden ogen soms als een bemost takje of een tak begroeid met korstmossen. Een klein aantal lijkt op bladeren, deze wandelende takken staan wel bekend als wandelende bladeren.[13] Bij de meeste soorten heeft het lichaam een glad oppervlak maar bij een aantal wandelende takken komen wratachtige uitsteeksels, bobbels of stekels voor op het lichaam.
Lengte en gewicht
De lichaamslengte varieert sterk; de soorten uit de geslachten Miniphasma en Grylloclonia blijven relatief klein. Mannetjes van Grylloclonia minima worden tot maximaal 1,7 centimeter lang. Van de soort Miniphasma prima is alleen een vrouwtje bekend, van 1,7 cm, de mannetjes zijn nog niet ontdekt maar blijven vermoedelijk nog iets kleiner.[14] Phobaeticus kirbyi, die in 1907 werd beschreven, bereikt een maximale lichaamslengte van 32,8 centimeter, exclusief poten. Deze soort wordt in de literatuur vaak vermeld als de langste soort, hoewel de soort Phobaeticus serratipes (ontdekt in 1835) een grotere lengte heeft als de poten worden meegerekend. In 2008 werd op Sabah (Borneo) Phobaeticus chani ontdekt, die net iets langer wordt. Exclusief poten meet het langst gevonden exemplaar 35,7 centimeter en met de poten erbij 56,7 cm.[15] In 2017 werd een exemplaar van de Chinese soort Phryganistria chinensis opgemeten met een totale lichaamslengte van 64 centimeter. Het is hiermee volgens het Guinness Book of Records het langste insect ter wereld.[16]
Wandelende takken staan ook bekend als de een van de zwaarlijvigste insecten. De zwaarst bekende wandelende tak is Heteropteryx dilatata, waarvan een zwanger vrouwtje meer dan 65 gram kan wegen. Alleen de krekel Deinacrida heteracantha wordt met 70 gram nog zwaarder.
Lichaamskleur
Wandelende takken hebben vrijwel altijd een groene of bruine lichaamskleur. Slechts enkele soorten hebben een gele, rode of blauwe kleur. Veel wandelende takken kunnen van kleur veranderen, afhankelijk van de omstandigheden. Een wandelende tak kan zijn lichaamskleur aanpassen aan de sterkte van het licht en ook de omgevingskleur speelt een rol.[17] De soort Sceptrophasma hispidulum heeft overdag een bleekgele kleur en krijgt 's nachts een donkere oranjerode kleur.[18]
Bij de veel bestudeerde Indische wandelende tak is deze kleurverandering goed onderzocht. De jonge nimfen verkrijgen hun uiteindelijke volwassen kleur deels door de kleur van de ondergrond waarop ze leven. Nimfen die op een lichte ondergrond verblijven worden groen en die op een donkere ondergrond worden uiteindelijk bruin. Exemplaren die als volwassene geheel groen zijn, blijken niet van kleur te kunnen veranderen. Bruin gekleurde dieren hebben vier soorten pigmentstoffen in de cellen van de huid: groene, bruine, oranje en gele. De bruine pigmenten kunnen zich op- en neer in de huidcellen verplaatsen en de oranje pigmenten kunnen zich in de cel verspreiden of samenklonteren. Wanneer de bruine pigmenten zich verplaatsen in de bovenste huidlaag en de oranje pigmenten worden samengetrokken wordt de kleur donkerder en vice versa. De groene en gele pigmenten kunnen zich echter niet verplaatsen.
De kleurverandering volgt een bepaald dag- en nachtritme op basis van de lichtintensiteit; overdag is het dier lichter dan 's nachts. Wanneer de wandelende tak bij wijze van proef overdag in het donker wordt gehouden en 's nachts aan licht wordt blootgesteld duurt het nog weken eer de kleurverandering zich weer heeft aangepast. Dit komt omdat het interval van de veranderende lichtintensiteit via de oogzenuwen in het centraal zenuwstelsel wordt 'opgeslagen'. De pigmentverdeling in de huid wordt waarschijnlijk bepaald door het periodiek uitscheiden van hormonen.[18]
Wandelende takken kunnen onder bepaalde omstandigheden polymorfie vertonen. Als ze in grote dichtheden voorkomen, veranderen ze drastisch van lichaamskleur, net zoals voorkomt bij treksprinkhanen. Deze specifieke vorm van polymorfie wordt kentromorfie genoemd. De normale gecamoufleerde vorm verandert bij dieren die in grote dichtheden leven in een felle schrikkleur.[19]
Geslachtsonderscheid
Mannetjes zijn in de regel aanzienlijk kleiner dan vrouwtjes. Vrouwtjes hebben met name een groter achterlijf, waarin ze de relatief grote eieren dragen. De kop en het maag-darmstelsel, die extra voedingstoffen moeten verzamelen voor de grote eieren, zijn eveneens groter dan bij mannetjes. Dit is ook het geval bij andere geleedpotigen zoals sprinkhanen en spinnen. Er zijn wel enkele uitzonderingen op deze regel; bij Eurycantha calcarata zijn mannetjes bijna net zo groot en bij de soort Oncotophasma martini zijn mannetjes zelfs groter dan vrouwtjes.
Mannetjes en vrouwtjes hebben vaak een verschillende lichaamskleur, zodat de geslachten hiermee kunnen worden onderscheiden. Bij de soort Heteropteryx dilatata hebben de mannen een bruine lichaamskleur, terwijl de vrouwen groen van kleur zijn. Over het algemeen zijn mannetjes ook veel beweeglijker en vaker actief dan de vrouwtjes.[17]
Kop

Pe = pedicel
Sc = scapus
O = oog
Pmx = maxillaire palp
Plb = labiale palp
Ma = mandibel
De kop van een wandelende tak is relatief klein en niet erg beweeglijk, en lijkt sterk op die van sprinkhanen. De kop is naar voren gericht en is meestal relatief smal maar sterk gewelfd. De ogen kunnen zowel klein als relatief groot zijn en hebben een ronde tot ovale vorm. Ze bestaan uit zogenaamde facetogen, dit is een complex van kleine lensjes die tezamen een beeld vormen van de omgeving. Bij een aantal soorten zijn aan de kop ook enkelvoudige lensoogjes of ocelli aanwezig. Ocelli komen alleen voor bij gevleugelde soorten.[20]
Aan de voorzijde van de kop zijn de mandibels of kaken en de maxillae of achterkaken gelegen aan de bovenzijde en het labrum of lip aan de onderzijde. Beide hebben een paar aanhangsels die dienen om het voedsel de mond in te manoeuvreren. Bij wandelende takken zijn deze aanhangsels langer en breder dan bij andere groepen van insecten. De monddelen hebben een bijtende en kauwende configuratie.
Aan de bovenzijde van de kop zijn de antennes of voelsprieten gepositioneerd. Deze kunnen zowel zeer lang en draadvormig zijn als juist zeer kort en nauwelijks zichtbaar. De meeste soorten hebben erg lange voelsprieten, die aan de kop zijn gehecht met de scapus, het breedste deel van de antenne. De scapus is het enige antennedeel dat voorzien is van spieren en is hierdoor het enige beweegbare deel. De scapus is middels een tweede antennelid, de pedicel, verbonden met de rest van de antenne. De pedicel is aanzienlijk korter dan de scapus. Het langste deel van de antenne bestaat uit het flagellum (vrij vertaald: zweep), dit zijn een aantal gelijkgevormde delen die naar het einde toe dunner worden. Het flagellum bestaat bij wandelende takken uit acht tot meer dan honderd segmenten.[20] Een uitzondering vormen de soorten uit het geslacht Paragongylopus, die slechts drie antenneleden bezitten. De antennes hebben zowel een tastzintuiglijke als een chemosensorische functie. Ze worden gebruikt om de omgeving af te speuren naar obstakels, maar kunnen ook bepaalde chemische stoffen waarnemen en temperatuursveranderingen vaststellen.
Veel soorten hebben opvallende stekels of andere structuren aan de kop, waarvan de functie nog niet precies bekend is. Bij een aantal soorten doen de vele stekels aan de bovenzijde van de kop denken aan een doornenkroon. Bactrododema tiaratum heeft twee oor-achtige uitsteeksels op de kop.
Borststuk
Het borststuk van insecten bestaat uit drie delen die borststuk- of thoraxsegmenten worden genoemd. Bij wandelende takken is het voorste deel relatief zeer kort en hiermee zijn ze te onderscheiden van veel andere langgerekte insecten zoals de langwerpige bidsprinkhanen uit het geslacht Thesprotia. Dit voorste segment van het borststuk wordt de prothorax genoemd en draagt het voorste paar poten. Aan de voorzijde van de prothorax is dorsaal het halsschild of pronotum gelegen. Bij sommige wandelende takken zijn aan weerszijden van het halsschild openingen van speciale afweerklieren gelegen, die gebruikt worden om vijanden af te schrikken. Het tweede en middelste deel van het borststuk, de mesothorax, wordt dat aan de bovenzijde beschermd door een verharde plaat, het scutum. De mesothorax draagt het middelste potenpaar en bij de gevleugelde soorten zijn hier de voorvleugels aangehecht. De mesothorax is verreweg het langste lichaamsdeel van de wandelende tak. Met name bij de gevleugelde soorten is de mesothorax langer en breder, omdat hierin de vleugelspieren zijn geborgen. Het achterste deel heet de metathorax. Ook dit deel kan relatief zeer lang zijn bij de staafvormige soorten die in grassen leven. De metathorax wordt aan de bovenzijde beschermd door een verharde plaat, het metanotum. De metathorax draagt het achterste potenpaar en bij gevleugelde soorten de achtervleugels. Dit achterste borststuksegment is versmolten met het eerste deel van het achterlijf.
Poten

De poten zijn ontwikkeld tot grote en langgerekte looppoten, waarmee de dieren zich verplaatsen tussen bladeren en takken. De poten reiken als ze naar achteren worden gestrekt in de regel veel verder dan de achterlijfspunt. Ze bezitten flexibele gewrichten en veel soorten kunnen hiermee wiegende bewegingen maken om een takje na te bootsen dat meebeweegt met de wind. Ondanks de lange poten kunnen wandelende takken niet snel rennen en ze kunnen ook niet springen.
De poten zijn met een ronde coxa aan het lichaam gehecht, gevolgd door de trochanter of dijbeenring. De coxa is relatief groot in vergelijking met die van andere insecten. De dijbeenring is smal en is het meest flexibele pootgewricht. De volgende segmenten bestaan uit het sterk verlengde femur of dij en de eveneens relatief lange tibia of scheen.
Het laatste deel van de poot bestaat uit de tarsus of voet. Deze is verdeeld in vijf verschillende leden, die de tarsomeren worden genoemd. Een uitzondering vormen de soorten uit de familie Timematodea, die slechts drie leden hebben. De eerste tarsomeer is aanmerkelijk langer dan de overige delen, de laatste zijn ongeveer even lang. Aan het eind van de tarsus is de pretarsus gelegen. Deze bevat gekromde, verharde haakjes, die de tarsale klauwen worden genoemd en kussenachtige structuren, de aurolia. Ook op het midden van de onderzijde van de eerste paar tarsomeren, gezien vanaf de pootbasis, zijn dergelijke kussentjes gelegen, de euplantulae. Soms is ook op het laatste tarsussegment een dergelijke structuur aanwezig.[21] De aurolia en euplantulae zijn kleine hechtkussentjes die een vergelijkbare functie hebben als de kleefvoetjes van sommige hagedissen, zoals gekko's. Ze zijn aan de onderzijde voorzien van kleine haartjes die het contactoppervlak vergroten. Hierdoor blijven de poten als het ware 'plakken' aan de ondergrond, maar kunnen ook gemakkelijk worden losgemaakt als de poot onder een bepaalde hoek van het oppervlak wordt getild.[22]
Vleugels
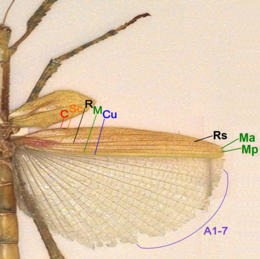
Sc = subcosta
R = radius
M = media
Cu = cubitus
Rs = radiaalsector
Ma = media anterior
Mp = media posterior
A = anaaladeren
Zowel de voorvleugels aan de bovenzijde als de achtervleugels aan de onderzijde hebben een verharde basis die doet denken aan de bouw van de vleugels van wantsen. Wandelende takken hebben in beginsel vleugels, maar bij zo'n zestig procent van alle soorten zijn de vleugels in de loop van de evolutie gereduceerd of helemaal verdwenen. Bij de gevleugelde soorten zijn het vaak alleen de mannetjes die kunnen vliegen. Daarentegen beschikken bij de soort Extatosoma tiaratum alleen de vrouwtjes over vleugels.[23] Bij andere soorten zoals Necroscia annulipes hebben alle exemplaren grote vleugels, waarmee ze zeer goed kunnen vliegen.
Veel van de gevleugelde soorten hebben ontwikkelde achtervleugels maar sterk gedrongen voorvleugels die nauwelijks als zodanig te herkennen zijn. Het vleugeloppervlak is bij deze soorten sterk verkleind en de vleugel is omgevormd tot een flapje. Zelfs de volledig gevleugelde soorten staan bekend als slechte vliegers en met name zwangere vrouwtjes kunnen door hun toegenomen gewicht niet meer opstijgen.[24] Bij een aantal soorten hebben alleen de mannetjes ontwikkelde vleugels waarmee ze kunnen vliegen en hebben de vrouwtjes rudimentaire vleugels die niet functioneel zijn. De vleugels zijn vaak doorzichtig en niet voorzien van kleuren, maar er zijn enkele uitzonderingen. De vleugels van wandelende bladeren zijn groen en de vleugeladeren zijn zo gerangschikt en gekleurd dat ze op bladnerven lijken.
De vleugels zijn te onderscheiden van die van andere insecten door de vorm en de positie van de vleugeladeren. De vleugelstructuur van wandelende takken lijkt nog het meest op die van wantsen, hoewel ze hieraan niet verwant zijn. De ader aan de bovenrand van de achtervleugel is onvertakt en wordt de costa genoemd, die ontbreekt op de voorvleugels. Hieronder is de tweede vleugelader, de subcosta, gelegen, die eveneens geen vertakkingen heeft. De derde ader is de radius. Deze heeft geen vertakkingen op de voorvleugels maar splitst zich wel op de achtervleugels. De vierde vleugelader is de media, die zich aan de vleugelbasis vertakt in de voorste (posterior) en de achterste (anterior) ader. De vijfde vleugelader is de cubitus en deze is weer onvertakt. De voorvleugels zijn voorzien van twee anale aders aan de onderzijde van de vleugel, de achtervleugels hebben er veel meer.
Achterlijf
Het achterlijf bestaat uit elf segmenten die terga (enkelvoud: tergum) worden genoemd. De achterlijfssegmenten zijn vaak lastig van elkaar te onderscheiden omdat ze samengesmolten lijken te zijn. Bij een aantal soorten hebben ze duidelijke knoop-achtige segmentgrenzen. Hierdoor lijkt het achterlijf op bepaalde plantendelen zoals de stengels van bamboeplanten, die eveneens verdikkingen hebben tussen de verschillende segmenten. De relatieve grootte van de verschillende achterlijfssegmenten kan een belangrijk determinatiekenmerk zijn. Bij de soort Hermarchus leytensis bijvoorbeeld, is het eerste achterlijfssegment twee keer zo lang als het metanotum, het tweede tot het vijfde deel zijn opvolgend steeds iets langer, terwijl segmenten zes tot acht juist steeds korter worden.[25]
Onder de laatste twee achterlijfssegmenten is bij de mannetjes de vomer aanwezig, die is te herkennen aan een verdikking aan de laatste twee achterlijfssegmenten. Deze langwerpige structuur past als een sleutel in een slot in twee kuiltjes aan achterlijf van het vrouwtje van dezelfde soort. Hierdoor kunnen mannetjes alleen met vrouwtjes van dezelfde soort paren en dit verkleint de kans op kruisingen. Vrouwtjes bezitten een ovipositor, die ook wel legboor of eiglijder wordt genoemd. Deze is al in het nimfstadium te zien en dient om de eieren de deponeren. De ovipositor is aan de laatste drie achterlijfssegmenten gepositioneerd.[26] Aan het laatste achterlijfssegment zijn de cerci gelegen, lichaamsaanhangsels die een tastzintuiglijke functie hebben. De cerci bestaan uit een enkel deel en zijn nooit geleed, zoals bij veel andere insecten voorkomt.
Het achterlijf bevat de voortplantingsorganen en het belangrijkste deel van de spijsverteringsorganen. Het spijsverteringsstelsel van wandelende takken is lange tijd onbegrepen geweest. Veel dieren die van planten leven hebben een symbiotische relatie met micro-organismen die de celwanden afbreken, zodat de voedingsstoffen door de darmen kunnen worden opgenomen. Aangezien wandelende takken sterke zuren en oxiderende spijsverteringssappen produceren was het maar de vraag of micro-organismen wel zouden overleven in het spijsverteringsstelsel. Uit recent onderzoek naar de spijsvertering blijkt dat de insecten zelf enzymen produceren om de celwanden te verteren en inderdaad geen gebruik maken van eencelligen. Het langwerpige en dunne achterlijf is ook niet geschikt om een dergelijk fermentatieproces te huisvesten.[27]
Onderscheid met andere insecten

Wandelende takken zijn met maar weinig andere insectengroepen te verwarren. Een van de groepen die een sterke gelijkenis vertonen met wandelende takken zijn de sprinkhanen uit de familie Proscopiidae. Ze hebben een sprieterige lichaamsvorm, een enigszins gelijkende lichaamsbouw en springpoten, hoewel ze die weinig gebruiken. Proscopiidae zijn daarnaast gemakkelijk te herkennen aan de langwerpige, opstaande kop, waarin de ogen helemaal bovenaan geplaatst zijn.
Een andere groep vormen de vleugelloze, op sprinkhanen gelijkende Mantophasmatodea. Deze vrij onbekende insecten uit zuidelijk Afrika hebben geen springpoten en hebben niet zo'n langwerpig lichaam als de meeste wandelende takken.
Daarnaast zijn verschillende soorten bidsprinkhanen lastig te onderscheiden van wandelende takken door hun sterk gecamoufleerde en langwerpige lichaam en lange poten. Bidsprinkhanen hebben een zeer beweeglijke kop en relatief grote ogen waaraan ze te herkennen zijn.
Sommige wantsen lijken op wandelende takken zoals de staafwants (Ranatra linearis) uit de familie waterschorpioenen (Nepidae). Deze wants leeft echter niet op het land maar in zoet water en bovendien is het voorste potenpaar omgebouwd tot grijppoten. De staafwants lijkt sprekend op een takje maar er zijn ook wantsen die ogen als bladeren.
Een aantal rechtvleugeligen uit de familie sabelsprinkhanen (Tettigonidae) lijkt sprekend op de bladeren van de planten waarop ze leven. Deze sprinkhanen worden wel aangeduid als bladsprinkhanen en worden in de Engelse taal leaf bugs genoemd. Deze insecten zijn met name te verwarren met wandelende bladeren.
Ten slotte zijn er soorten uit de onderorde van de cicaden die sterke gelijkenissen vertonen met plantendelen en verward kunnen worden met wandelende takken. Deze insecten zijn echter allemaal aanzienlijk kleiner; ze bereiken een lichaamslengte van hooguit enkele centimeters.
Voortplanting en ontwikkeling


Wandelende takken kunnen zich in tropische streken het gehele jaar door voortplanten en meerdere jaren oud worden. In meer gematigde streken zetten de vrouwtjes aan het eind van de zomer eieren af waarna ze spoedig sterven. De volgende generatie komt dan in de lente tevoorschijn waarna de cyclus opnieuw begint.
De voortplanting van wandelende takken kan op twee manieren gebeuren: maagdelijke voortplanting (parthenogenese) en geslachtelijke (seksuele) voortplanting. Sommige soorten kunnen zich alleen geslachtelijk voortplanten, andere soorten zowel geslachtelijk als (facultatief) ongeslachtelijk en bij weer andere soorten bestaan er überhaupt geen mannetjes en deze planten zich obligaat ongeslachtelijk voort.[4] Bij insectenkwekers wordt echter, indien beide manieren mogelijk zijn, seksuele voortplanting geprefereerd omdat de jongen sterker zijn en langer leven.
Geslachtelijke voortplanting
Ongeveer twee tot vier weken na de laatste vervelling worden de dieren geslachtsrijp en kunnen ze paren.[26] Bij wandelende takken blijven de vrouwtjes op hun voedselplant, terwijl de mannetjes actief op zoek gaan naar een partner. De vrouwtjes scheiden geurstoffen af om een mannetje te lokken. De mannetjes hebben een voorkeur voor kleinere vrouwtjes, omdat de kans groter is dat zij nog niet gepaard hebben. Ze zijn met name kieskeurig als er in de omgeving veel vrouwtjes aanwezig zijn. Mannetjes proberen te paren met meerdere vrouwtjes.[28]
Mannetjes zijn kleiner en lichter gebouwd dan vrouwtjes en soms wordt wel gedacht dat een vrouwtje haar 'jong' op de rug neemt. Aangezien wandelende takken geen enkele vorm van broedzorg kennen, betreft een dergelijk paartje steeds een koppeltje. Ook na de paring blijft het mannetje enige tijd op het vrouwtje zitten, dit dient niet zozeer om haar te bewaken tegen natuurlijke vijanden, zoals lange tijd werd gedacht, maar om te voorkomen dat andere mannetjes haar bevruchten. Voorafgaand aan de paring klimt een mannetje op het vrouwtje en maakt met zijn achterlijfspunt contact met die van haar. Hij blijft op haar zitten tot de paring voorbij is, dit kan enkele uren tot meerdere dagen duren. Het mannetje plakt een spermatofoor vast aan haar geslachtsopening, dit is een zaadhoudend pakketje. Het vrouwtje kan het sperma opslaan in een speciale holte in haar lichaam (spermatheek) en maanden later nog bevruchte eieren afzetten.
Maagdelijke voortplanting
Een opmerkelijk kenmerk van wandelende takken is het vermogen om zich maagdelijk voort te planten. Hierbij produceren de vrouwtjes onbevruchte eieren die desondanks wel uitkomen en voor levensvatbaar nageslacht zorgen. Bij maagdelijke voortplanting komen er geen mannetjes aan te pas.
Ongeslachtelijke voortplanting is in de regel niet voordelig voor een organisme omdat de nakomelingen altijd vrouwelijk zijn en er verhoudingsgewijs minder eieren uitkomen in vergelijking met bevruchte eitjes. De incubatietijd is in de regel ook aanmerkelijk langer. Er zijn hierop wel uitzonderingen, want bij soorten uit het geslacht Carausius is de incubatietijd en het uitkomstpercentage van onbevruchte en bevruchte eitjes ongeveer gelijk. Bij wandelende takken komen regelmatig gynandromorfen voor, dit zijn in wezen vrouwtjes die mannelijke eigenschappen hebben. Bij sommige parthenogene soorten komen bij een hogere temperatuur relatief veel exemplaren uit het ei die er mannelijk uitzien maar genetisch gezien toch vrouwelijk zijn. Ondanks paarpogingen kunnen ze geen levensvatbare eieren produceren.
Het vermogen tot maagdelijke voortplanting is waarschijnlijk pure noodzaak; wandelende takken verplaatsen zich zeer langzaam en bij de gevleugelde soorten zijn vrouwtjes te zwaar om te vliegen. Onder natuurlijke omstandigheden komen veel vrouwtjes simpelweg nooit een mannetje tegen en kunnen ze zo toch nakomelingen produceren.[24]
Van de Nieuw-Zeelandse soort Acanthoxyla prasina werd lange tijd aangenomen dat er helemaal geen mannetjes bestonden tot er in 2016 toch een gevonden werd. Dit mannetje werd aangetroffen in een geïntroduceerde populatie in het Engelse Cornwall en was waarschijnlijk het resultaat van een zeldzame mutatie.[29]
De mannetjes van de soort Clitarchus hookeri op Nieuw-Zeeland zijn even talrijk als de vrouwtjes. In Engeland echter, waar ze net als de voorgaande soort zijn geïntroduceerd, zijn mannetjes zeer zeldzaam of komen helemaal niet voor.[28]
Ei

De eieren van een wandelende tak zien er per soort iets anders uit, zodat op basis van de kenmerken van het ei de soort kan worden bepaald. Het aantal eieren varieert sterk per soort, de vrouwtjes van sommige soorten produceren vijftig eitjes gedurende hun gehele leven en anderen tot wel 1000. Een vrouwtje van de soort Hermarchus leytensis is met 2375 eieren gedurende haar leven de recordhoudster.[14] De grootste eieren zijn bekend van Haaniella echinata, ze hebben een ovale vorm en zijn tot twaalf millimeter lang en 0,3 gram zwaar. De langste eitjes zijn van de soort Medauromorpha regina, ze hebben een lengte tot zestien mm. Deze zijn echter zeer langwerpig van vorm en veel lichter dan die van de eerstgenoemde soort.
De frequentie waarmee de eieren worden afgezet verschilt sterk, van drie tot vijftien per nacht tot een enkel ei per week.[26] De incubatietijd loopt uiteen van enkele maanden tot een jaar. Dit is afhankelijk van de omgevingstemperatuur, bij de bekende Indische wandelende tak komen de eieren uit na vier tot zes maanden bij kamertemperatuur. Wordt de temperatuur verhoogd dan verschijnen de nimfen al na twee maanden en bij een onverwarmde ruimte pas na acht maanden. Sipyloidea sipylus uit Madagaskar kent een incubatieperiode van een maand bij een temperatuur van 24 tot 27 graden Celsius, maar bij lagere temperaturen kan het tot een jaar duren eer de eieren uitkomen.[30] Van soorten die in koelere streken leven is bekend dat de eieren in een ruststadium (diapauze) kunnen geraken en pas na drie jaar uitkomen.
Uiterlijk
De eieren zijn relatief groot in verhouding tot het lichaam van het vrouwtje. Ze hebben een harde schaal met aan de bovenzijde een dekseltje dat het operculum wordt genoemd. Het is aan de buitenzijde van het ei nog te zien waar het zaadje het ei is binnengekomen. Deze zogenaamde micropilar is gelegen op een vaak goed te onderscheiden deel van het ei dat bekend staat als de micropilarplaat. Deze plaat is vaak langgerekt tot ovaal of druppelvormig.[26]
De eieren zijn rond tot ovaal van vorm, slechts enkele soorten produceren langwerpige eieren. Eieren van de soort Phobaeticus chani hebben kenmerkende vleugelachtige uitsteeksels aan weerszijden, die waarschijnlijk dienen om het ei een stukje te kunnen laten zweven. Zo worden ze door de wind beter verspreid, vergelijkbaar met de gevleugelde vrucht van de esdoorn. Het ei kan de meest uiteenlopende vormen en oppervlakken hebben; die van de meeste soorten zijn goed gecamoufleerd en lijken op plantenzaden of uitwerpselen van insecten. Veel soorten produceren eieren die sprekend lijken op de zaden van hun voedselplant.
De vrouwtjes van Timema-soorten slikken bodemdeeltjes in, die in het lichaam aan het ei worden vastgeplakt als ze worden afgezet, zodat de eieren gecoat zijn met het bodemmateriaal waarop ze worden gedeponeerd.
Verspreiding
Wandelende takken maken geen nest, eicocon of spinselweb om de eitjes in op te bergen, maar de eieren worden een voor een afgezet. Tot 2015 werd gedacht dat dit voor alle soorten gold, maar in dat jaar werd in Vietnam een wandelende tak ontdekt die een eicocon of ootheca produceert. De wetenschappelijke naam van de soort is nog onbekend maar de wandelende tak behoort vrijwel zeker tot de onderfamilie Korinninae. Het dier zet een eicocon af met tientallen eieren die een voor een uitkomen. De cocon heeft een ovale vorm en wordt aan een blad gelijmd.[31]
De eieren van de wandelende tak worden op verschillende manieren verspreid. De meeste soorten laten de eieren simpelweg naar beneden vallen waarna ze op de bodem terecht komen. Het voordeel is dat het vrouwtje in de plant kan blijven zitten en niet op zoek hoeft naar een geschikte afzetplaats voor de eieren.[24] Bij Diapheromera femorata, die erg talrijk kan zijn, kan het voorkomen dat gedurende de voortplantingstijd een continu geruis te horen is, vergelijkbaar met een regenbui. Dit wordt veroorzaakt door de enorme aantallen eieren die de vrouwtjes continu op de bosbodem laten vallen.[14]
Veel wandelende takken hebben een manier gevonden om de eieren door mieren te laten beschermen en verspreiden. Deze soorten produceren eieren met een suikerhoudend goedje op het operculum. Dit wordt het capitulum genoemd en doet sterk denken aan een mierenbroodje of elaiosoom. Dit is een zoete afscheiding die sommige planten toevoegen aan hun zaden om mieren aan te trekken. De vettige, zoete substantie op het ei wordt erg gewaardeerd door de mieren, die het ei meenemen naar hun nest. Omdat de stoffen van het capitulum en het elaiosoom dezelfde chemische structuur hebben is het voor de mieren vrijwel onmogelijk om het verschil te bepalen.[24]
Het capitulum wordt geconsumeerd, terwijl het ei ongemoeid wordt gelaten en op de afvalhoop nabij het nest wordt gedeponeerd. Hier kan het embryo zich in alle rust verder ontwikkelen terwijl het door de mieren wordt beschermd tegen vijanden en parasieten die de eieren proberen binnen te dringen. Tevens biedt het mierennest enige bescherming tegen bosbranden.[20] Het trucje werkt niet bij alle mieren; van de soorten glimmende dikkop (Pheidole megacephala) en het spookdraaigatje (Tapinoma melanocephalum) is bekend dat ze de eieren van soorten uit het geslacht Graeffea juist opeten.[19]
Sommige wandelende takken verspreiden de eieren zelf en gebruiken het achterlijf om te eieren zo ver mogelijk van zich af te slingeren. Het ei kan zo enkele meters worden weggeworpen.[17] Voorbeelden hiervan zijn soorten uit het geslacht Extatosoma. Veel wandelende takken die in koelere gebieden leven zetten de eieren af in de bodem met de legboor. Dergelijke eieren hebben vaak een kogel-achtige vorm, waarbij het ei met de punt naar beneden in de bodem wordt gebracht om de weerstand te verminderen. Voorbeelden van soorten die de eieren in de bodem afzetten zijn die uit de geslachten Acacus, Centrophasma, Diesbachia en Orxines.[24] Soorten uit het geslacht Leptinia zetten de eieren af onder boomschors, zodat ze verborgen blijven. Vrouwtjes van de soort Timema californica plakken de eieren vast aan bladeren of schors of zelfs aan andere dieren.[26] Veel wandelende takken plakken de eieren los van elkaar aan bladeren, zoals de soorten uit de geslachten Sceptrophasma en Sipyloidea. Vertegenwoordigers van de geslachten Calvisia en Trachythorax zetten de eieren daarentegen in groepjes af.[24] Wandelende takken doen niet aan broedzorg, de jongen zijn nestvlieders en zijn direct zelfstandig.
Nimf

Wandelende takken behoren tot de groep van de Exopterygota en kennen een onvolledige gedaanteverwisseling. Dit betekent dat ze geen wormachtig larvestadium kennen, zoals bijvoorbeeld kevers, maar dat de jonge dieren al sterk op de ouderdieren lijken. De juvenielen worden bij deze insecten ook wel nimfen genoemd. Ze hebben nog niet alle lichaamskenmerken van hun ouders, zo zijn ze kleiner en ontbreken de geslachtsorganen en vleugels. Na iedere vervelling wordt de oude huid afgeworpen en komt een steeds grotere nimf tevoorschijn. Bij de meeste soorten vervellen de mannetjes vijf keer en de vrouwtjes zes keer voor ze geslachtsrijp zijn. Dit duurt afhankelijk van de soort en de omstandigheden ongeveer drie tot zes maanden.[26] Nimfen die bij een hogere temperatuur leven en veel voedsel voorhanden hebben, ontwikkelen zich aanmerkelijk sneller dan exemplaren die bij lagere temperaturen leven en over minder voedsel beschikken.[20]
Bij het vervellen wordt de oude huid afgeworpen en het hieruit kruipende exemplaar is een maatje groter geworden. Als het dier toe is aan een nieuwe huid blijft het enige tijd zitten en kromt het lichaam. De oude huid scheurt open op het halsschild, waarna eerst de prothorax tevoorschijn komt en vervolgens de kop en de voelsprieten. Daarna worden het borststuk en het achterlijf uit de oude huid getrokken. De achterlijfspunt blijft nog verbonden met de oude huid, terwijl de poten worden losgemaakt en ten slotte laat ook het achterste deel van de huid los. De wandelende tak zal enige tijd stil blijven zitten en kan door de nog zachte monddelen niet eten. Terwijl het dier de nieuwe huid laat uitharden, wordt de oude huid opgegeten om geen chitine – waaruit de huid is opgebouwd – verloren te laten gaan. Na enkele dagen is het nieuwe pantser volledig uitgehard.
Zowel tijdens als enige tijd na een vervelling is een wandelende tak zeer kwetsbaar voor vijanden, omdat het lichaam nog zacht is en de poten nog niet goed functioneren. Als het dier gestoord wordt tijdens een vervelling of in een ongunstige omgeving vervelt kunnen misvormingen ontstaan, wat fatale gevolgen kan hebben. Van in gevangenschap gehouden dieren is bekend dat een te hoge of juist te lage luchtvochtigheid kan leiden tot een mislukte vervelling, evenals een te kleine ruimte of te veel soortgenoten die voor verstoring zorgen.
Imago
Het volwassen insect wordt wel imago genoemd. De volwassen dieren zijn vaak tientallen tot honderden keren groter dan de net uit het ei gekropen nimfen. Bij de gevleugelde soorten hebben alleen de volwassen dieren ontwikkelde vleugels. Aangezien ze niet meer vervellen veranderen imago's niet meer van uiterlijk en afgebroken ledematen kunnen ook niet meer aangroeien.
De levensverwachting van wandelende takken varieert sterk per soort, mannetjes leven bovendien aanzienlijk korter dan vrouwtjes. Volwassen wandelende takken leven afhankelijk van de soort ongeveer drie maanden tot maximaal drie jaar.[26] Exemplaren in gevangenschap worden in de regel veel ouder dan dieren in het wild omdat ze niet te lijden hebben van natuurlijke vijanden, parasieten en voedselschaarste. De mannetjes van Phyllium bioculatum hebben waarschijnlijk de kortste levensverwachting en leven ongeveer drie weken. Exemplaren van de soort Dares verrucosus leven het langst en kunnen tot vijf jaar oud worden.[32]
Voedsel

Wandelende takken zijn zonder uitzondering planteneters (fytofaag) en leven van verschillende typen planten, variërend van bomen en struiken tot grassen. Ze eten uitsluitend het blad van de plant en zuigen geen plantensappen op, in tegenstelling tot het veel andere insecten. Het is de enige insectenorde waarvan alle leden uitsluitend leven van bladeren, een voedselspecialisatie die ook wel aangeduid wordt met folivoor of bladetend. De meeste soorten eten van bedektzadige planten, een kleiner aantal van naaktzadigen of van varens. De nimfen eten dezelfde voedselplant(en) als de volwassen dieren.[4]
Alle soorten hebben de neiging om zich van de bodem af te bewegen, dit wordt wel negatieve geotaxie genoemd. Hierdoor klimmen ze naar het hoogste deel van een plant, waar de jongste blaadjes groeien.[19] Wandelende takken leven daardoor meestal op enige hoogte in bomen of hoge struiken maar er zijn er ook die dichter bij de bodem leven op grassen en kruidachtige planten. Gevleugelde soorten zijn in staat zich snel te verspreiden en zijn hierdoor vaak voedselspecialisten die slechts van enkele soorten planten eten. Voorbeelden zijn de vaak volledig gevleugelde soorten uit de onderfamilie Necrosciinae. Soorten die niet kunnen vliegen zijn noodgedwongen polyfaag en leven van veel verschillende planten.[24]
Wandelende takken kunnen veel schade aanrichten aan een plant, zie ook onder het kopje plaaginsect. Ze staan bekend als bijzonder vraatzuchtig en zijn daarnaast relatief inefficiënte eters. Veel soorten eten niet het hele blad op maar knagen een stuk af waarna de rest van het blad op de grond valt en verloren gaat.
Vijanden
Wandelende takken vormen een eiwitrijke voedselbron en er zijn veel dieren die op ze jagen. Voorbeelden zijn vogels, reptielen en rovende geleedpotigen zoals spinnen. Ook vleermuizen zijn een belangrijke vijand, die de wandelende tak opsporen door middel van hun uitstekende gehoor. Het knagen aan planten brengt minieme geluidjes voort die door de vleermuis kunnen worden gedetecteerd en de wandelende tak heeft hierdoor niets aan zijn uitstekende camouflage.[33]

Geleedpotige vijanden zijn schorpioenvliegen en ook van bidsprinkhanen en mieren is bekend dat ze wandelende takken eten. Ze worden geparasiteerd door vele verschillende ongewervelde dieren. Bekende voorbeelden zijn de tweevleugeligen uit de familie sluipvliegen (Tachinidae). Deze parasiteren voornamelijk op de nimfen, de soort Euhallidaya sverinii tast daarentegen de eieren aan. Phasmophaga antennalis heeft er zelfs zijn naam aan te danken; Phasmo-phaga betekent vrij vertaald 'wandelende tak-etend'. Belangrijke vliesvleugelige parasieten van de eieren zijn verschillende soorten goudwespen, met name uit de onderfamilies Amiseginae en Loboscelidiinae, en wespen uit de familie Eupelmidae. Deze soorten zoeken gericht naar de eieren, waar ze hun eigen ei inbrengen en de uitsluipende larve vervolgens het embryo opeet. Van parasitisme is feitelijk geen sprake omdat het embryo van de wandelende tak hierbij niet wordt aangetast maar ten gronde gaat.
Als echte parasieten kunnen nematoden worden vermeld die de nimfen en imago's aantasten, zoals de nematoden uit de familie Mermithidae.[19] Deze zogenaamde paardenhaarwormen verlaten het lichaam van het geïnfecteerde dier als ze volgroeid zijn en zijn dan veel langer dan hun slachtoffer. Uit een mannetje van de soort Haaniella echinata, die maximaal acht centimeter lang wordt, verscheen eens een worm van maar liefst 54 cm lang.[34]
Van verschillende soorten bloedzuigende mijten is bekend dat ze wandelende takken aantasten door hun lichaamssappen op te zuigen. Mijten uit het geslacht Lepus zuigen bloed van de soort Anisomorpha buprestoides en de mijt Allidosoma froggatti doet dat bij de soort Podacanthus wilkinsoni. Een dergelijk bloedzuigend gedrag komt ook voor bij kleine muggen die tot de familie knutten (Ceratopogonidae) behoren.
Verdediging

Verreweg de meeste wandelende takken lijken op plantendelen en gaan volledig op in hun natuurlijke omgeving, wat ook wel crypsis wordt genoemd. Ook hun nachtactieve levenswijze helpt ze om niet op te vallen, want overdag blijven ze doodstil zitten op de plant waarin ze leven. Veel soorten eten pas als er enige wind staat, zodat ze minder opvallen. De meeste soorten blijven altijd op hun voedselplant, maar er zijn ook wandelende takken die de plant verlaten als het licht wordt en zich overdag verschuilen in de strooisellaag.
Wandelende takken blijven vaak op hun plaats als ze worden bedreigd, want ze kunnen niet snel wegrennen bij gevaar. Geen enkele soort kan springen, zoals wel bekend is van op hen gelijkende sprinkhanen. Veel soorten laten zich bij verstoring op de grond vallen en verstijven alsof ze dood zijn, waarbij de lange poten tegen het lichaam worden gehouden. Dit wordt ook wel schijndood genoemd en deze vorm van schijndood staat bekend als thanatose. Het verstijven van de spieren wordt aangeduid met catalepsie. Van een aantal soorten is bekend dat ze autotomie vertonen: als ze worden vastgepakt laten ze de desbetreffende poot los om zelf te kunnen ontsnappen. Net zoals bij hagedissen, die hun staart kunnen loslaten, breekt de poot op een vaste plaats af, tussen de trochanter (dijbeenring) en het femur (dij). De dieren hebben hier weinig last van en gaan op vijf poten verder. Bij jongere dieren groeit de poot na een aantal vervellingen weer geheel aan. Ook afgebroken of afgebeten antennes kunnen weer aangroeien.
Wandelende takken kunnen het lichaam laten schommelen als het waait, zodat het net lijkt of het lichaam meebeweegt met de plant waarop ze zitten. Dit versterkt de camouflage, omdat de meeste insecten zich juist zo stil mogelijk proberen te houden als ze worden verstoord. Het schommelen heeft ook een visuele functie omdat het helpt bij het inschatten van de afstand tot objecten in de omgeving.
Een andere techniek die wordt toegepast is het gebruik van schrikkleuren, bijvoorbeeld door de Indische wandelende tak (Carausius morosus). Hierbij houden de dieren hun poten of vleugels in rust bij elkaar, maar als ze worden verstoord worden de poten of de vleugels gespreid waarbij de felle kleuren aan de binnenzijde van de poten tevoorschijn komen. Hierdoor worden roofdieren even afgeleid, waarna de wandelende tak zich laat vallen en het roofdier het insect niet meer kan vinden tussen de echte takjes op de grond.
Sommige wandelende takken lijken op andere dieren waar hun natuurlijke vijanden liever bij uit de buurt blijven. De nimfen van Extatosoma tiaratum lijken op stekende mieren, de volwassen exemplaren van de soorten uit het geslacht Eurycantha hebben een sterke gelijkenis met schorpioenen. Ze krommen het achterlijf voorwaarts boven het lichaam, wat doet denken aan de staart van een schorpioen. Een dergelijke nabootsing van gevaarlijke dieren door onschuldige imitaties komt veel voor bij insecten en wordt mimicry genoemd.

Andere wandelende takken hebben fel gekleurde achtervleugels, die in rust onder de gecamoufleerde voorvleugels worden gehouden maar bij bedreiging plotseling worden getoond. Peruphasma schultei spreidt bij verstoring de kleine achtervleugels om de lichaamscontouren te vergroten, waarna deze snel weer snel worden ingeklapt. Roofdieren raken hierdoor verward, omdat ze de locatie van hun prooi zo moeilijk kunnen lokaliseren.
Een soort die dreiggedrag vertoont is Heteropteryx dilatata, de vrouwtjes kunnen hun vleugels namelijk tegen elkaar strijken wat een luid sissend geluid veroorzaakt. Dit wordt striduleren genoemd. De stridulatieorganen zijn relatief slecht ontwikkeld in vergelijking met bijvoorbeeld sprinkhanen.
Van een aantal wandelende takken is bekend dat ze actief de aanval openen op een vijand. Soorten uit de geslachten Eurycantha en Heteropteryx kunnen met de stekels op hun poten verwondingen veroorzaken als ze worden vastgepakt. Heteropteryx dilatata verdedigt zich met behulp van stekels op de poten, met name die aan de dij en de scheen. De wandelende tak schopt met de achterpoten met grote kracht achterwaarts waardoor de stekels de huid doorboren. De stekels zijn lang, scherp en achterwaarts gekromd. Een rake klap is ook voor een mensenhand een pijnlijke ervaring aangezien de stekels tot het bot kunnen doordringen.[14]
Soorten die giftige of onsmakelijke planten eten, zoals Peruphasma schultei, gebruiken een irriterende vloeistof om roofdieren weg te jagen. Ze regurgiteren het laatst opgenomen voedsel in de hoop dat een predator met het braaksel in aanraking komt en de aanval staakt. Andere soorten kennen reflexbloed, zoals ook bekend is van onder andere lieveheersbeestjes. Bij bedreiging persen ze lichaamsvloeistoffen (hemolymfe) uit de gewrichten tussen de poten. Deze afweerstoffen hebben een zeer smerige geur en smaak. Reflexbloeden is bekend van de vertegenwoordigers van de geslachten Eurycantha, Orxines en Sipyloidea.
De hoogst ontwikkelde vorm van verdediging vindt men bij de soorten die een gerichte straal vloeistof op een belager spuiten om deze te verjagen. Er zijn ook wandelende takken die een straal afweerstoffen over zichzelf spuiten om zich zo onsmakelijk te maken. Van de Papoea-Nieuw-Guinese soort Megacrania nigrosulfurea is bekend dat beide methoden worden gebruikt.[35]
De meest extreme vorm van een dergelijke verdediging kennen de soorten uit het geslacht Anisomorpha. Deze wandelende takken spuiten zuur uit een gepaarde opening in het prothorax. Het zuur belemmert de ademhaling van de aanvaller, smaakt en ruikt zeer smerig en kan als het in de slijmvliezen zoals de ogen terecht komt voor ernstige irritatie zorgen.[26] Het wordt wel vergeleken met traangas en is zowel effectief tegen gewervelde als ongewervelde dieren.[4]
Relatie met de mens
Wandelende takken zijn bij mensen op grote schaal populair als huisdier en in sommige delen van de wereld worden ze gezien als voedsel. Ze kunnen echter ook aanzienlijke schade veroorzaken aan gewassen. Een groot aantal soorten wordt door menselijke activiteiten bedreigd.
Bedreigingen door de mens

Veel soorten worden bedreigd door de mens, met name de landbouwmethode 'hakken en branden' vormt een gevaar.
De soort Carausius scotti wordt bedreigd doordat de geïntroduceerde ceylonkaneelboom (Cinnamomum verum) het natuurlijke verspreidingsgebied overwoekert. De belangrijkste waardplant is de nestvaren (Asplenium nidus) maar ook andere varens waarvan de soort leeft worden hierdoor weggeconcurreerd. Het gevolg is dat de wandelende tak over een steeds kleiner verspreidingsgebied beschikt omdat de voedselplanten verdwijnen.[36]
Door de internationale natuurbeschermingsorganisatie IUCN is aan een relatief klein aantal soorten (104) een beschermingsstatus toegewezen. Van de soorten worden er 62 als 'veilig' (Least Concern of LC) beschouwd, 23 als 'onzeker' (Data Deficient of DD), 2 soorten als 'kwetsbaar' (Vulnerable of VU), 10 als 'gevoelig' (Near Threatened of NT), 1 als 'bedreigd' (Endangered of EN) en 5 soorten staan te boek als 'ernstig bedreigd' (Critically Endangered of CR).[12]
De Peruaanse soort Peruphasma schultei heeft een zeer klein verspreidingsgebied maar omdat deze soort door de zwarte kleur, rode vleugels en goudgele ogen erg populair is in de handel in exotische dieren zijn er veel ex situ populaties. Hierdoor kunnen de dieren worden uitgezet in de natuur om de soort voor uitsterven te behoeden.
Een van de zeldzaamste wandelende takken, en tevens een van de meest bedreigde insecten, is Dryococelus australis. Deze soort kwam voor op het ten oosten van Australië gelegen Lord Howe-eiland, maar was al sinds 1920 niet meer gezien en werd beschouwd als uitgestorven. In 2001 werd deze wandelende tak herontdekt op het onbewoonde eilandje Ball's Pyramid. Het verdwijnen van deze soort op Lord Howe-eiland was het gevolg van de introductie van de zwarte rat (Rattus rattus) in het natuurlijke verspreidingsgebied. Veel andere soorten die leven in geïsoleerde gebieden als eilanden, worden eveneens bedreigd door de rat, zoals Cornicandovia australica[37] en Davidrentzia valida.[38]
De soort Pseudobactricia ridleyi wordt beschouwd als uitgestorven. Deze wandelende tak is slechts bekend van een enkel exemplaar uit Singapore. De soort is ondanks intensieve speurtochten in Singapore en naburige landen al meer dan honderd jaar niet meer gezien en de bossen waarin het dier leefde zijn verdwenen.[39]
Wandelende takken als voedsel en medicijn
Wandelende takken worden door sommige inheemse volken beschouwd als voedselbron. De bevolking van Papoea-Nieuw-Guinea eet exemplaren van Extatosoma popa of soorten uit het geslacht Eurycantha. Deze worden bij de oogst van bladeren gevangen en aan een stokje gespietst waarna ze al draaiend worden geroosterd tot de poten loslaten en vervolgens worden ze opgegeten. Van de Bidayuh uit Sarawak is beschreven dat ze eieren van Haaniella grayi verzamelen om op te eten. Hiertoe worden zwangere vrouwtjes gezocht die worden ontdaan van hun ongeveer 10 millimeter lange eieren. De eitjes worden eerst een halve minuut gekookt en vervolgens gepeld voor consumptie. De eieren zijn rijk aan proteïnen en daarnaast wordt geloofd dat ze helpen tegen diarree.[14] De Iban uit dezelfde regio eten exemplaren van Haaniella echinata.[32]
Eurycantha calcarata wordt in Papoea-Nieuw-Guinea gebruikt om vissen te vangen. Deze soort heeft sterk gekromde stekels aan de achterpoten die van het dier worden afgebroken en worden gebruikt als vishaak.[40]
De uitwerpselen van de soort Eurycnema versirubra worden in delen van China en Maleisië gebruikt om een thee te maken die klachten als astma en spierpijn zouden verlichten. De soort Megacrania nigrosulfurea wordt door de lokale bevolking van Papoea-Nieuw-Guinea als desinfectant gebruikt bij huidinfecties.[35]
Plaaginsect

Verschillende soorten zijn sinds de jaren 80 van de 19e eeuw bekend als plaaginsecten in delen van Noord-Amerika, China en Australië. Als ze in grote aantallen voorkomen kunnen wandelende takken enorme schade aanrichten aan gewassen door de bladeren op te eten. De vraatschade is dan vergelijkbaar met die van kevers, zoals de eveneens bladetende coloradokever, die aardappelplanten aantast, en met treksprinkhanen, die in Afrika grote verwoestingen aan de oogst kunnen toebrengen.
Ongevleugelde soorten verspreiden zich veel langzamer dan gevleugelde wandelende takken. Van de soort Diapheromera femorata is bekend dat de soort zich ongeveer 200 meter per jaar kan verplaatsen.[19] De bestrijding van wandelende takken beperkt zich vaak tot het gebruik van insecticiden, ook worden wel delen van het bos in brand gezet om ze uit te roken. In delen van China wordt de schimmel Beauveria bassiana ingezet om ze te bestrijden als ze een plaag vormen.
Bepaalde soorten die in Australië leven kunnen massaal voorkomen en een eucalyptus-boom geheel kaalvreten. Ook soorten die gehele bossen aantasten zijn bekend. In vochtige delen van Australië komen verschillende soorten voor die naast de natuurlijke begroeiing ook landbouwgewassen als voedsel gebruiken. Dergelijke wandelende takken worden gezien als erg schadelijk voor de voedselvoorziening van de mens.
De Noord-Amerikaanse soort Diapheromera femorata is in staat om in korte tijd een eik geheel te ontbladeren.[41] Bekende gewassen die door de mens worden geteeld en kunnen lijden door vraatschade zijn onder andere mango (Mangifera indica), kokospalm (Cocos nucifera), robinia (Robinia pseudoacacia), hartbladige haagbeuk (Carpinus cordata), guave (Psidium guajava), geurende schroefpalm (Pandanus tectorius), zwarte peper (Piper nigrum), maïs (Zea mays), zure kers (Prunus cerasus), pruim (Prunus domestica), Kaapse jasmijn (Gardenia jasminoides) en aardappel (Solanum tuberosum).[19]
Huisdier

Wandelende takken horen tot de weinige insecten die populair zijn als exotisch huisdier. Ze worden gehouden in een terrarium, dat zowel uit glas als plastic kan bestaan. Een belangrijke regel is dat het terrarium minstens twee tot drie keer de grootte van de volwassen wandelende takken moet hebben. De insecten hebben dit nodig om goed te kunnen vervellen, anders kunnen er misvormingen optreden waaraan ze kunnen overlijden.
De meest gehouden soort is de Indische wandelende tak (Carausius morosus), die tot acht centimeter lang kan worden. Vrijwel alle andere soorten die in gevangenschap worden gehouden behoren tot de volgende geslachten; Acrophylla, Anisomorpha, Aplopus, Baculkum, Calynda, Clonopsis, Creoxylus, Dares, Dyme, Epidares, Eurycantha, Extatosoma, Haaniella, Hesperophasma, Leptynia, Libethra, Lonchodes, Neohirasea, Oreophoetes, Orxines, Paramenexenus, Phyllium en Rhaphiderus.
De gewenste temperatuur en relatieve vochtigheid in het terrarium hangen af van de plaats van herkomst van de wandelende takken. De omstandigheden daar kunnen sterk van elkaar verschillen. Het terrarium moet enige ventilatie hebben om beschimmeling te voorkomen. De openingen mogen niet te groot zijn omdat de nimfen gemakkelijk ontsnappen. Als bodembedekking wordt meestal kranten- of keukenpapier gebruikt, aangezien dit goedkoop is. Om het terrarium er mooier uit te laten zien kan eventueel zand, potgrond of een andere bodembedekker voor terrariums toegepast worden.
De meeste soorten die in gevangenschap worden gehouden eten braam en klimop, maar sommige soorten zijn kieskeurig wat hun voeding betreft en lusten alleen moeilijker te verkrijgen planten zoals Eucalyptus. Wandelende takken drinken weinig of niets en halen het vocht uit de bladeren. Het is daarom belangrijk uitdroging van de terrariumplanten te voorkomen. Indien er te weinig voeding in het terrarium te vinden is, bestaat de kans dat wandelende takken uit honger aan soortgenoten gaan knagen.
Studie en indeling van de wandelende takken

Wandelende takken worden door veel particulieren als huisdier gehouden vanwege de relatief eenvoudige verzorging en het opvallende uiterlijk van de dieren. Daarnaast planten ze zich in gevangenschap relatief gemakkelijk voort.
Phasmid Study Group
Liefhebbers van wandelende takken en bladeren hebben zich verenigd in de Phasmid Study Group. Dit is een collectief met leden variërend van amateurentomologen, professionele schrijvers en wetenschappers tot wereldwijd erkende experts op het gebied van wandelende takken. Het doel van de groep is het bestuderen, het houden en grootbrengen, en het zich laten voortplanten van wandelende takken in gevangenschap op een zo natuurlijk mogelijke manier. Nieuwe informatie wordt openbaar gedeeld op de website. Veel van de informatie over wandelende takken is ontsloten door de PSG. Van ruim zeventig wandelende takken is de wetenschappelijke soortaanduiding vernoemd naar een van de leden van deze groep.[42]

De groep heeft iedere soort wandelende tak een zogeheten PSG-nummer gegeven, waardoor taxonomische veranderingen minder verwarring veroorzaken. Het PSG-nummer verandert namelijk niet als bijvoorbeeld de geslachtsnaam van een soort op basis van nieuwe inzichten wordt aangepast. Dit laatste is onder meer gebeurd met de soorten uit het geslacht Baculum, die later tot het geslacht Ramulus werden gerekend, maar hun PSG-nummer hebben behouden.
Onderlinge indeling
In november 2022 telde de 'Phasmida Species File' ongeveer 3500 verschillende soorten wandelende takken.[43] Het totale aantal, dus inclusief de nog niet ontdekte soorten, is waarschijnlijk aanzienlijk hoger. In het wild komen soorten voor die nog niet wetenschappelijk zijn beschreven en ook zijn nog niet alle exemplaren onderzocht die in museumcollecties zijn opgenomen. Een moeilijkheid bij de classificatie van wandelende takken is dat de dieren zich in verschillende delen van de wereld steeds opnieuw hebben moeten aanpassen aan veranderende micro-habitats. De gelijkende uiterlijke kenmerken van verschillende soorten zeggen hierdoor weinig over hun gemeenschappelijke voorouders.[24]
Lange tijd werden de wandelende takken verdeeld in twee onderordes op basis van de aan- of afwezigheid van een structuur aan de schenen van de poten, de area apicalis. Deze indeling in grofweg twee groepen, de Anareolatae (die de structuur wel bezitten) en de Areolatae (zonder deze aanpassing), bleek niet monotypisch en wordt beschouwd als verouderd.[44]
Een andere indeling splitst de wandelende takken in drie groepen; de Agathemerodea (vertegenwoordigd door een enkel geslacht), de Timematodea (eveneens een geslacht) en de Verophasmatodea (alle andere geslachten). Op basis van moleculair onderzoek vallen de Agathemerodea waarschijnlijk binnen de Verophasmatodea, reden waarom deze indeling niet breed wordt gedragen.[45]
De insecten worden door de Phasmid Study Group verdeeld in twee onderordes en veertien families. Deze indeling is gebaseerd op het aantal segmenten aan de uiteinden van de poten. Dertien van deze families worden tot de onderorde Euphasmatodea gerekend, vroeger wel Verophasmatodea genoemd, en worden verder verdeeld in vier superfamilies. De veertiende familie, Timematidae, wordt tot een aparte onderorde gerekend; Timematodea. Er is binnen deze groep slechts een enkel geslacht, met veertien soorten. De reden van deze afsplitsing is dat het een zeer primitieve familie betreft. De vertegenwoordigers van deze familie blijven klein, tot ongeveer 2,5 centimeter, en hebben drieledige uiteinden van de poten (tarsi) in plaats van vijfledige, zoals alle andere wandelende takken. Daarnaast hebben alle mannetjes een vergrote lob aan hun rechter-cercus, dit terwijl asymmetrische lichaamsdelen over het algemeen zeer ongebruikelijk zijn bij insecten.
Een nieuwe classificatie verdeelt de wandelende takken in twee groepen: de Oriophasmata (de 'oostelijke' wandelende takken) en de Occidophasmata (de 'westelijke' wandelende takken).[13] Deze in 2019 geïntroduceerde indeling wordt echter nog niet breed erkend.
Evolutie

De evolutie van wandelende takken is lastig te bestuderen, omdat fossielen heel zeldzaam zijn, vooral van complete exemplaren en de eieren. Vrijwel alle bekende fossielen bestaan slechts uit afdrukken van vleugels die op het sediment zijn achtergebleven. Lange tijd werd gedacht dat de meeste fossielen van wandelende takken uit het Paleozoïcum stammen, tot bleek dat de fossielen die aan de groep werden toegedicht afdrukken waren van andere insecten. De oudste fossielen waarvan zeker is dat het wandelende takken betreft, zijn afkomstig van de familie Hagiphasmatidae en dateren uit het Krijt. Uit het latere Eoceen zijn fossielen bekend van ongeveer 44 tot 49 miljoen jaar oud, die werden gevonden in zowel de Messelgroeve in Duitsland als in de John Day Fossil Beds National Monument in Oregon, Verenigde Staten. Jongere fossielen zijn bekend uit het Oligoceen en het Mioceen en zijn aangetroffen in schalielagen in de Florissant Fossil Beds National Monument in Colorado, VS. Ook in Baltisch barnsteen uit Scandinavië en Dominicaans barnsteen zijn wandelende takken in dezelfde periode gefossiliseerd.[4] Er zijn alleen fossielen bekend van de grootste onderorde Euphasmatodea, van de Timematodea zijn nog geen fossielen gevonden.
Wandelende takken moeten volgens genetische analyses veel ouder zijn. De twee onderordes splitsten zich, volgens een researchartikel uit 2019, af aan het begin van het Krijt, ongeveer 120 miljoen jaar geleden.[46] Volgens een artikel uit 2022 vond deze splitsing zelfs nog eerder plaats, tussen 190 en 270 miljoen jaar geleden.[47] Aan het einde van het Krijt vond een tweede splitsing plaats tussen de geslachten Orthomeria en Abrosoma en alle andere moderne soorten. Gedurende het Paleogeen en het Neogeen vonden nog vele afsplitsingen plaats, die hebben geleid tot de verschillende moderne families. De onderlinge verwantschappen tussen de families zijn echter nog grotendeels onbekend.
In 2013 werd een fossiel van de Palaeochresmoda grauvogeli gevonden, een insect dat ongeveer 245 miljoen jaar geleden leefde in het Trias. Het werd aangetroffen in de Franse plaats Hangviller en lijkt sterk op een wandelende tak of een voorloper ervan. De groep waartoe het dier behoort is niet met zekerheid bekend.[48]
Wandelende bladeren werden lange tijd gezien als een vroege aftakking van de wandelende takken, maar tegenwoordig vermoeden biologen dat ze juist een recentere oorsprong hebben.[13] Het oudst bekende wandelend blad is Eophyllium messelensis; een fossiel van deze soort werd eveneens gevonden in de Messelgroeve en de soort is ernaar vernoemd. De Eophyllium messelensis leek al sterk op de huidige wandelende bladeren, maar had nog geen verbrede voorpoten, zoals voorkomt bij veel van de moderne vertegenwoordigers.[49]
Schema: indeling van de Polyneoptera
Wandelende takken behoren tot de insecten uit het cohort Polyneoptera, die onder andere ook de Zoraptera, de oorwormen, de steenvliegen en de rechtvleugeligen omvat. De Polyneoptera is verder verdeeld in twee zustergroepen: de kakkerlakachtigen of Dictyoptera (de bidsprinkhanen en de kakkerlakken en termieten) en de Mecynoptera. Deze laatste is onderverdeeld in de Notoptera die de Grylloblattodea en Mantophasmatodea omvat en de Eukinolabia waartoe alle webspinners en wandelende takken behoren.[46]
Onderstaand is een schema weergegeven van de positie van de wandelende takken binnen de groepen van insecten die het sterkst aan ze verwant zijn.

| Polyneoptera |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Lijst van families van wandelende takken
Onderstaand is een overzicht weergegeven van de twee onderorden van wandelende takken, met de vier superfamilies en veertien families, hun onderlinge rangschikking en de auteur, het verspreidingsgebied en een voorbeeldsoort. Uitgestorven superfamilies, zoals Susumanioidea, zijn niet opgenomen.
| Familie | Indeling | Verspreiding | Voorbeeldsoort | Afbeelding voorbeeldsoort |
|---|---|---|---|---|
| Onderorde Euphasmatodea | ||||
| Superfamilie Aschiphasmatoidea Brunner von Wattenwyl, 1893 | ||||
| Familie Aschiphasmatidae Brunner von Wattenwyl, 1893 |
93 soorten in 20 geslachten, 1 onderfamilie | Tropische delen van zuidoostelijk Azië. | Aschiphasma annulipes |  |
| Familie Damasippoididae Zompro, 2004 |
Zes soorten in twee geslachten. | Afrika; Madagaskar | Damasippoides albomarginatus | |
| Familie Prisopodidae Brunner von Wattenwyl, 1893 |
54 soorten in vijf geslachten, twee onderfamilies. | Zuidelijk Noord-Amerika, Zuid-Amerika, uiterst zuidelijk Afrika, Azië | Melophasma antillarum |  |
| Superfamilie Bacilloidea Brunner von Wattenwyl, 1893 | ||||
| Familie Anisacanthidae Günther, 1953 |
33 soorten in 11 geslachten, drie onderfamilies. | Afrika; Madagaskar | Parectatosoma mocquerysi | |
| Familie Bacillidae Brunner von Wattenwyl, 1893 |
54 soorten in 19 geslachten, drie onderfamilies. | Europa, Afrika | Europese wandelende tak (Bacillus rossius) |
 |
| Familie Heteropterygidae Kirby, 1896 |
133 soorten in 26 geslachten, drie onderfamilies. | Australië, Azië | Hoploclonia abercrombiei |  |
| Superfamilie Phyllioidea Brunner von Wattenwyl, 1893 | ||||
| Familie Phylliidae Brunner von Wattenwyl, 1893 |
103 soorten in 10 geslachten, één onderfamilie. | Australië, Azië | Phyllium bioculatum |  |
| Superfamilie Pseudophasmatoidea Rehn, 1904 | ||||
| Familie Agathemeridae Bradler, 2003 |
Acht soorten in 1 geslacht, één onderfamilie. | Zuid-Amerika; Argentinië en Chili | Agathemera maculafulgens |  |
| Familie Heteronemiidae Rehn, 1904 |
67 soorten in 14 geslachten, een onderfamilie. | Noord-Amerika en Zuid-Amerika, Afrika; Madagaskar | Hoploclonia cuspidata |  |
| Familie Pseudophasmatidae Rehn, 1904 |
308 soorten in 47 geslachten, drie onderfamilies. | Noord-Amerika en Zuid-Amerika, Afrika, Europa, Australië, Azië, Oceanië. | Pseudophasma acanthonotus |  |
| Familie Diapheromeridae Kirby, 1904 |
608 soorten in 73 geslachten, drie onderfamilies. | Wereldwijd | Hermagoras hosei |  |
| Familie Lonchodidae Brunner von Wattenwyl, 1893 |
1138 soorten in 159 geslachten, twee onderfamilies. | Zuid-Amerika, Europa, zuidelijk Afrika, Australië, Azië, Oceanië. | Phenacephorus cornucervi |  |
| Familie Phasmatidae Leach, 1815 |
679 soorten in 126 geslachten, acht onderfamilies. | Wereldwijd, uitgezonderd Europa en Noord-Amerika | Phobaeticus serratipes |  |
| Onderorde Timematodea Kevan, 1982 | ||||
| Familie Timematidae Caudell, 1903 |
21 soorten in een geslacht en één onderfamilie. | Westelijk Noord-Amerika | Timema californicum |  |
Externe links
- (nl) – Verzorgingsfiches en foto's van wandelende takken en wandelende bladeren
- (nl) – Wandelende takken en wandelende bladeren houden als huisdier
- (en) – Phasmid Study Group
Bronvermelding
| Dit artikel is op 16 november 2022 in deze versie opgenomen in de etalage. |