Чулни орган


| Сензорски нервни систем | |
|---|---|
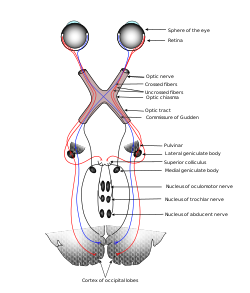 Типични сензорски систем: видни систем, илустрован класичном Грејовом сликом 722 – Ова шема приказује проток информација од очију до централних веза оптичких нерва и трактова, до визуелног кортекса. Област В1 је регион мозга који учествује у виду. | |
| Детаљи | |
| Идентификатори | |
| Латински | organa sensuum |
| TA | A15.0.00.000 |
| FMA | 78499 75259, 78499 |
| Анатомска терминологија | |
Чулни органи или сензорни нервни систем су део нервног система који је одговоран за обраду сензорских информација. Сензорски систем се састоји од сензорских неурона (укључујући сензорске рецепторске ћелије), неуронске путеве, и делове мозга који учествују у сензорској перцепцији. Најзаступљенији сензорски системи су они за вид, слух, додир, укус, мирис, и баланс. Укратко, чула су трансдуктори из физичког света у царство ума, где се информација интерпретира и формира перцепција света око нас.[1]
Рецептивно поље је област тела или окружења на коју рецепторски орган и ћелијски рецептори одговарају. На пример, део света које око може да види, је рецептивно поље ока; светлост коју сваки штапић или купа може да види, је њихово рецептивно поље.[2] Рецептивна поља се могу идентификовати за визуелни систем, аудиторни систем и соматосензорски систем.
У састав чула кичмењака улазе три типа чулних елемената:
- примарне чулне ћелије – настају прве у еволуцији на површини тела; имају способност примања и преношења надражаја; задржале су се у чулу вида и мириса код кичмењака;
- слободни нервни завршеци – ћелије које леже дубље испод површине тела и са њом су у вези преко наставака;
- секундарне чулне ћелије – имају само способност примања, а не и преношења надражаја; присутне су у већини чула кичмењака.
Према врсти дражи коју примају чула се деле на:
- механичка – примају дражи додира, притиска а такође и звучне и друге таласе;
- хемијска – примају дражи различитих хемијских материја у гасовитом и течном стању;
- оптичка – примају светлосне дражи, чуло вида
Механичка чула
* тактилни органи (чуло додира) – најраспрострањеније и еволуционо најстарије чуло; примају дражи из спољашње средине и унутрашњих органа; представљени су или слободним нервним завршецима или секундарним чулним ћелијама; код кичмењака постоје тактилна телашца која реагују на притисак, температуру, бол;
- бочни органи – код водених кичмењака; налазе се на глави и дуж бочних линија тела; примају надражаје покрета и струјања воде, помоћу њих се оријентишу у кретању кроз воду (заобилажење предмета);
- инфундибуларни орган – постоји код амфиоксуса и риба; налази се на доњој страни међумозга; служи за оријентацију риба у погледу дубине воде;води порекло од предњег отвора нервне цеви.
- статички (равнотежни) и слушни органи – обједињени су у заједнички стато-акустички апарат
Органи чула слуха и равнотеже
Смештени су у унутрашњем уху. У њему се налазе полукружни каналићи (чуло равнотеже) и пуж (чуло слуха). Полукружни каналићи и пуж су утопљени у течности (перилимфа) и испуњени су течношћу (ендолимфа). У ендолимфи полукружних каналића су кристали калцијум-карбоната који надражују чулне ћелије. У пужу је смештен Кортијев орган чије чулне ћелије примају треперења перилимфе.
Код гмизаваца, птица и сисара су се развили и помоћни делови који примају и преносе звучна треперења – спољашње и средње ухо. Спољашње ухо се састоји од ушне шкољке и спољашњег слушног канала који се завршава бубном опном. Средње ухо обухвата бубну дупљу са бубном опном и три слушне кошчице. Чекић (malleus) је причвршћен за бубну опну, а за њега је покретно везан наковањ (incus). Између средњег и унутрашњег уха је овално окно на које належе узенгија (stapes). Звучна треперења изазивају треперења бубне опне, па се преко слушних кошчица и овалног окна преносе на перилимфу.
Чуло вида
Очи кичмењака припадају мехурастом типу и могу бити:
- непарне очи – има их мали број кичмењака и то као темено (колоусте, неке рибе и гуштери) и пинеално око (код колоуста, а код осталих кичмењака је претворено у епифизу);
- парне очи.
Парне очи
Изграђене су по типу затвореног очног мехура (очна јабучица). Очна јабучица обавијена је са три опне:
- беоњачом – спољашња опна, која на предњем крају образује провидну рожњачу;
- судовњачом – богата крвним судовима; на предњем крају образује дужицу (различито обојена) која на средини има отвор – зеницу; иза дужице лежи очно сочиво;
- мрежњачом (ретина) –унутрашња опна; она је најсложенији део ока; у њој се налазе чулне ћелије:
- штапићи – одговорни за разликовање светлости и таме; код сисара су бројнији од чепића;
- чепићи – одговорни за уочавање боја и детаља предмета; највише их има у средини мрежњаче (на линији која пролази кроз центар ока) на месту које се зове жута мрља – место најоштријег вида.
На месту где очни нерв излази из ока нема чулних елемената па се то место назива слепа мрља.
Хемијска чула
Припадају им чуло мириса и укуса.
Чуло мириса (олфактивни органи)
Смештено је у парним носним (мирисним) јамама. Код копнених кичмењака мирисне јаме добијају и функцију спровођења ваздуха до плућа, па се образује носно-ждрелни канал који повезује мирисне органе са усном дупљом.
Чуло укуса
Представљено је групама секундарних чулних ћелија које заједно са потпорним ћелијама образују тзв. густативне квржице. Код риба су ове квржице распоређене по усној дупљи, ждрелу па чак и у кожи на површини тела. Код сувоземних кичмењака налазе се само на језику и непцима, а код сисара само на језику.
Постоје 4 основна осећаја укуса:
- сланог,
- слатког,
- горког и
- киселог.
Највећа осетљивост за поједине надражаје везана је за одређене зоне језика – врхом језика се осећа слатко, а задњим крајем горко итд. Не постоји чулна зона за љуто, већ се такве дражи преносе преко нервних завршетака као бол.

Чула и рецептори
Специфични број чула није усаглашен међу неуролозима услед различитих дефиниција у погледу тога шта сачињава чуло. Сидарта Гаутама и Аристотел класификују пет ‘традиционалних’ људских чула, што је постало широко прихваћено: додир, укус, мирис, вид, и слух. Друга чула која су добро позната код већине сисара, укључујући људе, обухватају ноцицепцију,[3] чуло равнотеже,[4][5][6][7][8] проприоцепцију,[9][10][11][12] и термоцепцију. Осим тога, показано је да неке нехумане животиње поседују алтернативна чула, укључујући магнетоцепцију[13][14][15][16] и електроцепцију.[17][18][19]
Рецептори
Иницијализација сензације потиче од одговора одређеног рецептора на физички стимулус. Рецептори који реагују на стимулус и иницирају процес сензације обично се групишу у четири различите категорије: хеморецептори, фоторецептори, механорецептори, и терморецептори. Сви рецептори примају својствене физичке стимулусе и преображавају сигнал у електрични акциони потенцијал. Овај акциони потенцијал затим путује дуж узлазних нервних влакана до специфичних можданих региона где бива обрађен и интерпретиран.[20]
Хеморецептори
Хеморецептори, или хемосензори, детектују одређене хемијске стимулусе и преображавају тај сигнал у електрични акциони потенцијал. Два примарна типа хеморецептора су:[21][22][23][24]
- Даљински хеморецептори који су интегрални при примању стимулуса у олфакторном систему путем олфакторних рецепторских неурона и неурона у Јакобсоновом органу.
- Директни хеморецептори који обухватају папиле укуса у густаторном систему, као и рецепторе у аортним телима који детектују промене концентрације кисеоника.[25]
Фоторецептори
Фоторецептори имају способност фототрансдукције, што је процес у коме се конвертује светлост (електромагнетско зрачење) у, поред осталих типова енергије, мембрански потенцијал.[26][27][28] Три примарна типа фоторецептора су:
- Чепићи су фоторецептори који прозводе знатан респонс на боју. Код људи три различита типа чешће кореспондирају примарном респонсу на кратке таласне дужине (плаво), средње таласне дужине (зелено), и дуге таласне дужине (жуто/црвено).[29]
- Штапићи су фоторецептори који су веома сензитивни на јачину светлости, те омогућава вид при слабом осветљењу. Концентрација и однос чепића и штапића су зависни од тога да ли је животиња диурнална или ноктурна. Код људи број штапића надмашује број чепића са односом од око 20:1, док код ноктурних животиња, као што је шумска сова, однос је око 1000:1.[29]
- Ганглијске ћелије ретине су присутне у адреналној медули и ретини, где оне учествују у симпатичком респонсу. Од око 1,3 милиона ганглионских ћелија присутних у ретини, сматра се да су 1-2% фотосензитивне ганглије.[30] Ове фотосензитивне ганглије учествују у свесном виду код неких животиња,[31] и сматра се да то чине и код људи.[32]
Механорецептори
Механорецептори су сензорни рецептори који одговарају на механичке силе, као што је притисак или дисторзија.[33][34] Док су механорецептори присутни у ћелијама длачица и играју интегралну улогу у вестибуларном и аудиторном систему, већина механорецептора се налази у кожи.[35][36] Они се групишу у четири категорије:
- Споро прилагођавајући рецептори типа 1 имају мала рецептивна поља и одговарају на статичку стимулацију. Ови рецептори првенствено долазе до изражаја при сензацијама облика и храпавости.[37]
- Споро прилагођавајући рецептори типа 2 имају велика рецептивна поља и одговарају на растезање. Слично типу 1, они производе непрекидне одговоре на континуиране стимулусе.[38]
- Брзо прилагођавајући рецептори имају мала рецептивна поља и у основи су перцепције клизања.[39][40]
- Пакинијски рецептори имају велика рецептивна поља и они су предоминантни рецептори за високо фреквентне вибрације.[41][39][42]
Терморецептори
Терморецептори су сензорни рецептори који одговарају на промене температуре.[43][44] Мада механизам путем кога ови рецептори делују није јасан, недавна открића су показала да сисари имају најмање два различита типа терморецептора:[45]
- Краусова телашца, или булбоидне корпускуле,[46] који детектују температуре изнад телесне температуре.
- Руфанијева телашца детектују температуре испод телесне температуре.[47][48][49][50]
Ноцицептори
Ноцицептори одговарају на потенцијално оштећујуће стимулусе слањем сигнала у кичмену мождину и мозак.[51][52][53][54] Овај процес, који се назива наноцепција, обично узрокује перцепцију бола.[55][56][57] Они су присутни у унутрашњим органима, као и на површини тела.[58] Ноцицептори откривају различите врсте оштећујућих стимулуса или стварна оштећења.[59] Они који реагују само када су ткива оштећена, познати су као „спавајући” или „неми” ноцицептори.
- Термални ноцицептори се активирају излагањем шкодљивој топлоти или хладноћи на разним температура.
- Механички ноцицептори одговарају на прекомерни притисак или механичке деформације.
- Хемијски ноцицептори одговарају на широк варијетет хемикалија, неке од којих су знаци оштећења ткива. Они учествују у детекцији појединих зачина у храни.
Сензорни кортекс
Сви стимулуси које примају горенаведени рецептори се преносе до акционог потенцијала.[60][61][62] Он се преноси дуж једног или више аферентних неурона према специфичној области мозга.[63][64][65] Док се термини сензорни кортекс често неформално користи за референциње соматосензорног кортекса, термин се прецизније односи на вишеструке области мозга у којима долази до пријема чулних инпута ради обраде. За пет традиционалних чула код људи, тиме су обухваћени примарни и секундарни кортекси разних чула: соматосензорски кортекс, визуелни кортекс, аудиторни кортекс, примарни мирисни кортекс, и кортекс укуса.[66] Други модалитети исто тако имају кореспондирајуће сензорне кортексе, укључујући вестибуларни кортекс за осећај равнотеже.[67][68][69][70][71]

Соматосензорски кортекс
Лоциран у теменом можданом режњу, примарни соматосензорски кортекс је примарно прихватно подручје за сигнале чула додира и проприоцепције[9][72][73] у соматосензорном систему.[74] Овај кортекс се даље дели у Бродманова поља 1, 2, и 3. Бродманово поље 3 се сматра примарним центром за обраду информација сематосензорног кортекса, јер оно прима знатно већу количину података од таламуса, садржи неуроне који су високо респонсивни на сематосензорне стимулусе, и може да евоцира соматске сензације кроз електричну стимулацију. Поља 1 и 2 примају највећи део свог улаза из поља 3. Исто тако постоје путеви за проприоцепцију (преко малог мозга), и моторну контролу (преко Бродмановог поља 4). Такође погледајте: С2 Секундарни соматосензорски кортекс.
Визуелни кортекс
Визуелни кортекс се односи на примарни визуелни кортекс, означен са В1 или Бродманово поље 17, као и на екстрастријатна визуелно кортексна поља В2-В5.[75] Лоциран у потиљачном режњу, В1 делује као примарна релејна станица за визуелни инпут, која преноси информације на две примарне путање означене као дорсални и вентрални ток. Дорсални ток обухвата поља В2 и В5, и користи се за интерпретацију визуелног „где” и „како”. Вентрални ток обухвата поља В2 и В4, и користи се у интерпретацији „шта” аспекта.[76] Повећања у активности негативних задатака су уочена у вентралној мрежи пажње, након наглих промена у сензорним стимулусима,[77] на почетку и офсету блокова задатака,[78] и на крају завршеног покушаја.[79]
Аудиторни кортекс
Лоциран у слепоочном можданом режњу,[80][81] аудиторни кортекс је примарно рецептивно подручје за звучне информације.[82][83][84] Аудиторни кортекс се састоји од Бродманових поља 41 и 42, такође познатих као предње попречно темпорално поље 41 и задње попречно темпорално поље 42, респективно.[85][86] Оба поља делују на сличан начин и интегрална су у пријему и обради сигнала трансмитованих из аудиторних рецептора.[87][88]
Примарни мирисни кортекс
Лоциран у слепоочном можданом режњу, примарни олфакторни кортекс је примарна рецептивна област за мирис, или њух.[89][90][91] Јединствено за олфакторне и густаторне системе, бар код сисара, је имплементација периферних и централних механизама дејства. Периферни механизми обухватају олфакторне рецепторске неуроне[92] који преносе хемијске сигнале дуж мирисних живаца, који се завршавају у олфакторној луковици.[93][94] Хеморецептори који учествују у каскади олфакторних нерва користе Г протеин спрегнуте рецепторе за слање својих хемијских сигнала низ поменуту каскаду.[95][96][97] Централни механизми обухватају конвергенцију олфакторних нервних аксона у гломерули у олфакторној луковици,[98] где се сигнал затим трансмитује до предњег олфакторског језгра, пириформног кортекса, медијалне амигдале, и енториналног кортекса, сви од којих сачињавају примарни олфакторни кортекс.
За разлику од вида и слуха, олфакторне луковице нису унакрсно хемисферне; десна луковица је повезана са десном хемисфером и лева луковица са левом хемисфером.

Кортекс укуса
Кортекс укуса је примарна рецептивна област за укус. Реч укус се користи у техничком смислу за специфично упућивање на сензације које потичу од папила укуса на језику. Пет форми укуса које детектује језик обухватају киселост, горчину, слаткост, сланост и укус протеина, звани умами. За разлику од тога, термин арома се односи на доживљај створен путем интеграције укуса са мирисом и тактилном информацијом. Густаторни кортекс се састоји од две примарне структуре: предње инсуле, лоциране на инсуларном режњу, и фронталног оперкулума, лоцираног на чеоном режњу. Слично олфакторном кортексу, густаторна путања оперише путем периферног и централног механизма. Периферни рецептори укуса, лоцирани на језику, меком непцу, ждрелу, и једњаку, преносе примљени сигнал до примарних сензорних аксона, где се сигнал пројектује на језгро самотарног тракта у продуженој мождини, или на густаторно језгро комплекса самотарног тракта. Сигнал се затим трансмитује до таламуса, који пројектује сигнал на неколико региона неокортекса, укључујући густаторни кортекс.[99]
На неуронску обраду укуса утичу у готово свакој фази обраде истовремене соматосензорне информације са језика, тј. осећај текстуре. Мирис, за разлику од тога, се не комбинује са укусом ради стварање ароме све до виших региона кортикалне обраде, као што су инсула и орбитофронтални кортекс.[100]
Види још
- Мултисензорска интеграција
- Неуронска адаптација
- Неуронско кодирање
- Сензор
Референце
- ^ Krantz, John. „Experiencing Sensation and Perception - Chapter 1: What is Sensation and Perception?” (Pdf). стр. 1.6. Приступљено 16. 5. 2013.
- ^ Kolb & Whishaw: Fundamentals of Human Neuropsychology (2003)
- ^ Craig, A D (2003). „A new view of pain as a homeostatic emotion”. Trends in Neurosciences. 26 (6): 303—7. PMID 12798599. S2CID 19794544. doi:10.1016/S0166-2236(03)00123-1.
- ^ Seeley, R., VanPutte, C., Regan, J., & Russo, A. (2011). Seeley's Anatomy & Physiology (9th ed.). New York, NY: McGraw Hill
- ^ Albertine, Kurt. Barron’s Anatomy Flash Cards
- ^ "How Does Our Sense of Balance Work?" How Does Our Sense of Balance Work?U.S. National Library of Medicine, 12 Jan. 2012. Web. 28 Mar. 2016.
- ^ "Semicircular Canals." Semicircular Canals Function, Definition & Anatomy. Healthline Medical Team, 26 Jan. 2015. Web. 28 Mar. 2016.
- ^ Tillotson, Joanne. McCann, Stephanie. Kaplan’s Medical Flashcards. Apr. 02. 2013.
- ^ а б Gandevia, Simon; Proske, Uwe (1. 9. 2016). „Proprioception: The Sense Within”. The Scientist. Приступљено 25. 7. 2018.
- ^ Fox, Richard; Barnes, Robert D.; Ruppert, Edward E. (2003). Invertebrate Zoology: A Functional Evolutionary Approach (на језику: Anglais) (7th изд.). Australia, Brazil, Japan, Korea: Brooks/Cole. ISBN 9780030259821.
- ^ Bastien, Renaud; Bohr, Tomas; Moulia, Bruno; Douady, Stéphane (8. 1. 2013). „Unifying model of shoot gravitropism reveals proprioception as a central feature of posture control in plants”. Proceedings of the National Academy of Sciences. 110 (2): 755—760. Bibcode:2013PNAS..110..755B. ISSN 0027-8424. PMC 3545775
 . PMID 23236182. doi:10.1073/pnas.1214301109 .
. PMID 23236182. doi:10.1073/pnas.1214301109 .
- ^ Hamant, Olivier; Moulia, Bruno (1. 10. 2016). „How do plants read their own shapes?”. New Phytologist (на језику: енглески). 212 (2): 333—337. ISSN 1469-8137. PMID 27532273. doi:10.1111/nph.14143.
- ^ Wiltschko, F.R.; Wiltschko, W. (2012). „Chapter 8 - Magnetoreception”. Ур.: Carlos López-Larrea. Sensing in Nature. Advances in Experimental Medicine and Biology. 739. Springer. ISBN 978-1-4614-1703-3. S2CID 41131723. doi:10.1007/978-1-4614-1704-0. Непознати параметар
|name-list-style=игнорисан (помоћ) - ^ Foley, Lauren E.; Gegear, Robert J.; Reppert, Steven M. (2011). „Human cryptochrome exhibits light-dependent magnetosensitivity”. Nature Communications. 2 (Article 356): 356. Bibcode:2011NatCo...2E.356F. PMC 3128388 . PMID 21694704. doi:10.1038/ncomms1364.
- ^ Wiltschko, Wolfgang; Wiltschko, Roswitha (2008). „Magnetic orientation and magnetoreception in birds and other animals”. Journal of Comparative Physiology A. 191 (8): 675—93. PMID 15886990. S2CID 206960525. doi:10.1007/s00359-005-0627-7.
- ^ Blakemore, R. (1975). „Magnetotactic Bacteria”. Science. 190 (4212): 377—379. Bibcode:1975Sci...190..377B. PMID 170679. S2CID 5139699. doi:10.1126/science.170679.
- ^ Hofle, M., Hauck, M., Engel, A. K., & Senkowski, D. (2010). Pain processing in multisensory environments. [Article]. Neuroforum, 16(2), 172.
- ^ Heiligenberg, Walter (1977) Principles of Electrolocation and Jamming Avoidance in Electric Fish: A Neuroethological Approach Springer-Verlag. Heiligenberg, Walter (1977). Principles of Electrolocation and Jamming Avoidance in Electric Fish: A Neuroethological Approach. Springer. ISBN 9780387083674.
- ^ Lewicki, Michael S.; Olshausen, Bruno A.; Surlykke, Annemarie; Moss, Cynthia F. (2014). „Scene analysis in the natural environment”. Frontiers in Psychology. 5: 199. PMC 3978336 . PMID 24744740. doi:10.3389/fpsyg.2014.00199 .
- ^ Sensory receptor, From: A Dictionary of Psychology Архивирано 2009-01-12 на сајту Wayback Machine
- ^ Rawson, Nancy E.; Yee, Karen K. (2006). „Transduction and Coding”. Taste and Smell. Advances in Oto-Rhino-Laryngology. 63. стр. 23—43. ISBN 3-8055-8123-8. PMID 16733331. doi:10.1159/000093749.
- ^ Saunders, CJ; Christensen, M; Finger, TE; Tizzano, M (2014). „Cholinergic neurotransmission links solitary chemosensory cells to nasal inflammation”. Proceedings of the National Academy of Sciences. 111 (16): 6075—80. Bibcode:2014PNAS..111.6075S. PMC 4000837 . PMID 24711432. doi:10.1073/pnas.1402251111 .
- ^ Finger, Thomas E.; Kinnamon, Sue C. (1. 1. 2013). „A taste for ATP: neurotransmission in taste buds”. Frontiers in Cellular Neuroscience. 7: 264. PMC 3866518 . PMID 24385952. doi:10.3389/fncel.2013.00264 .
- ^ Nurse, Colin A. (2013). „Signal processing at mammalian carotid body chemoreceptors”. Semin Cell Dev Biol. 24 (1): 22—30. PMID 23022231. doi:10.1016/j.semcdb.2012.09.006.
- ^ Satir, P. & Christensen, S.T. (2008) Structure and function of mammalian cilia. in Histochemistry and Cell Biology, Vol 129:6
- ^ Foster, R.G.; Provencio, I.; Hudson, D.; Fiske, S.; Grip, W.; Menaker, M. (1991). „Circadian photoreception in the retinally degenerate mouse (rd/rd)”. Journal of Comparative Physiology A. 169 (1): 39—50. PMID 1941717. S2CID 1124159. doi:10.1007/BF00198171.
- ^ Hecht, S.; Shlar, S.; Pirenne, M.H. (1942). „Energy, Quanta, and Vision”. Journal of General Physiology. 25 (6): 819—840. PMC 2142545 . PMID 19873316. doi:10.1085/jgp.25.6.819.
- ^ Baylor, D.A.; Lamb, T.D.; Yau, K.W. (1979). „Responses of retinal rods to single photons”. The Journal of Physiology. 288: 613—634. PMC 1281447 . PMID 112243. doi:10.1113/jphysiol.1979.sp012716.
- ^ а б "eye, human." Encyclopædia Britannica. Encyclopædia Britannica Ultimate Reference Suite. Chicago: Encyclopædia Britannica, 2010.
- ^ Foster, R. G.; Provencio, I.; Hudson, D.; Fiske, S.; De Grip, W.; Menaker, M. (1991). „Circadian photoreception in the retinally degenerate mouse (Rd/Rd)”. Journal of Comparative Physiology A. 169 (1): 39—50. PMID 1941717. S2CID 1124159. doi:10.1007/BF00198171.
- ^ Jennifer LE, Olivia ND, Kwoon YW, Nazia MA, Shih-Kuo C, Tara L, Jordan MR, Glen TP, David MB, Samer H (2010). „Melanopsin-Expressing Retinal Ganglion-Cell Photoreceptors: Cellular Diversity and Role in Pattern Vision”. Neuron. 67 (1): 49—60. PMC 2904318 . PMID 20624591. doi:10.1016/j.neuron.2010.05.023.
- ^ Horiguchi, H.; Winawer, J.; Dougherty, R. F.; Wandell, B. A. (2012). „Human trichromacy revisited”. Proceedings of the National Academy of Sciences. 110 (3): E260—E269. ISSN 0027-8424. PMC 3549098 . PMID 23256158. doi:10.1073/pnas.1214240110 .
- ^ Winter, R.; Harrar, V.; Gozdzik, M.; Harris, L. R. (2008). „The relative timing of active and passive touch. [Proceedings Paper]”. Brain Research. 1242: 54—58. PMID 18634764. S2CID 11179917. doi:10.1016/j.brainres.2008.06.090.
- ^ Adrian ED, Umrath K (октобар 1929). „The impulse discharge from the pacinian corpuscle”. The Journal of Physiology. 68 (2): 139—54. PMC 1402853 . PMID 16994055. doi:10.1113/jphysiol.1929.sp002601.
- ^ Johansson RS, Flanagan JR (мај 2009). „Coding and use of tactile signals from the fingertips in object manipulation tasks” (PDF). Nature Reviews. Neuroscience. 10 (5): 345—59. PMID 19352402. S2CID 17298704. doi:10.1038/nrn2621.
- ^ McNulty PA, Macefield VG (децембар 2001). „Modulation of ongoing EMG by different classes of low-threshold mechanoreceptors in the human hand”. The Journal of Physiology. 537 (Pt 3): 1021—32. PMC 2278990 . PMID 11744774. doi:10.1111/j.1469-7793.2001.01021.x.
- ^ Johnson KO, Hsiao SS (1992). „Neural mechanisms of tactual form and texture perception”. Annual Review of Neuroscience. 15: 227—50. PMID 1575442. doi:10.1146/annurev.ne.15.030192.001303.
- ^ Torebjörk HE, Ochoa JL (децембар 1980). „Specific sensations evoked by activity in single identified sensory units in man”. Acta Physiologica Scandinavica. 110 (4): 445—7. PMID 7234450. doi:10.1111/j.1748-1716.1980.tb06695.x.
- ^ а б Talbot WH, Darian-Smith I, Kornhuber HH, Mountcastle VB (март 1968). „The sense of flutter-vibration: comparison of the human capacity with response patterns of mechanoreceptive afferents from the monkey hand”. Journal of Neurophysiology. 31 (2): 301—34. PMID 4972033. doi:10.1152/jn.1968.31.2.301.
- ^ Johansson RS, Westling G (1987). „Signals in tactile afferents from the fingers eliciting adaptive motor responses during precision grip”. Experimental Brain Research. 66 (1): 141—54. PMID 3582528. S2CID 22450227. doi:10.1007/BF00236210.
- ^ Biswas A, Manivannan M, Srinivasan MA (2015). „Multiscale layered biomechanical model of the pacinian corpuscle”. IEEE Transactions on Haptics. 8 (1): 31—42. PMID 25398182. S2CID 24658742. doi:10.1109/TOH.2014.2369416.
- ^ Biswas A, Manivannan M, Srinivasan MA (2015). „Vibrotactile sensitivity threshold: nonlinear stochastic mechanotransduction model of the Pacinian Corpuscle”. IEEE Transactions on Haptics. 8 (1): 102—13. PMID 25398183. S2CID 15326972. doi:10.1109/TOH.2014.2369422.
- ^ Darian-Smith, Ian; Johnson, KO; LaMotte, C.; Y, Shigenaga; Kenins, P.; P, Champness (1979). „Warm fibers innervating palmar and digital skin of the monkey: responses to thermal stimuli.”. Journal of Neurophysiology. 42 (5): 1297—1315. PMID 114608. doi:10.1152/jn.1979.42.5.1297.
- ^ Viana, Felix; E, la Peña; Belmonte, C. (2002). „Specificity of cold thermotransduction is determined by differential ionic channel expression.”. Nature Neuroscience. 5 (3): 254—260. PMID 11836533. S2CID 21291629. doi:10.1038/nn809.
- ^ Krantz, John. Experiencing Sensation and Perception Архивирано на сајту Wayback Machine (17. новембар 2017). Pearson Education, Limited, 2009. p. 12.3
- ^ W. Krause. Die terminalen Körperchen der einfach sensiblen Nerven. Hannover, 1860.
- ^ „8”. Ganong's Review of Medical Physiology (23rd изд.). TATA McGraw-Hill Lange. 2010. стр. 150. ISBN 978-0-07-067722-7.
- ^ Mountcastle, Vernon C. (2005). The Sensory Hand: Neural Mechanisms of Somatic Sensation. Harvard University Press. стр. 34.
- ^ „8”. Ganong's Review of Medical Physiology (23rd изд.). TATA McGraw-Hill Lange. 2010. стр. 150. ISBN 978-0-07-067722-7.
- ^ Guyton, Arthur c. E. Hall; John. „47”. Guyton & Hall Pocket Companion to Textbook of Medical Physiology (10 изд.). стр. 362. ISBN 978-81-8147-057-7.
- ^ „Lorimer Moseley on What is Pain?”. Архивирано из оригинала 04. 08. 2018. г. Приступљено 01. 11. 2018.
- ^ „NOI - Neuro Orthopaedic Institute”. www.noigroup.com. Архивирано из оригинала 17. 10. 2018. г. Приступљено 1. 11. 2018.
- ^ „Nociception and pain: What is the difference and why does it matter? - Massage St. Louis, St. Louis, MO”. www.massage-stlouis.com. Архивирано из оригинала 01. 11. 2018. г. Приступљено 01. 11. 2018.
- ^ Animals, National Research Council (US) Committee on Recognition and Alleviation of Pain in Laboratory (8. 12. 2017). Mechanisms of Pain. National Academies Press (US) — преко www.ncbi.nlm.nih.gov.
- ^ Sherrington C. The Integrative Action of the Nervous System. Oxford: Oxford University Press; 1906.
- ^ Portenoy, Russell K.; Brennan, Michael J. (1994). „Chronic Pain Management”. Ур.: Good, David C.; Couch, James R. Handbook of Neurorehabilitation. Informa Healthcare. ISBN 978-0-8247-8822-3.
- ^ „Assessing Pain and Distress: A Veterinary Behaviorist's Perspective by Kathryn Bayne”. Definition of Pain and Distress and Reporting Requirements for Laboratory Animals. Proceedings of the Workshop Held June 22, 2000. 2000.
- ^ Purves, D. (2001). „Nociceptors”. Ур.: Sunderland, MA. Neuroscience. Sinauer Associates.
- ^ Feinstein, B.; Langton, J.; Jameson, R.; Schiller, F. (1954). „Experiments on pain referred from deep somatic tissues”. J Bone Joint Surg. 36—A (5): 981—97. PMID 13211692. doi:10.2106/00004623-195436050-00007. Архивирано из оригинала 12. 2. 2008. г. Приступљено 6. 1. 2007.
- ^ Hodgkin AL, Huxley AF (1952). „A quantitative description of membrane current and its application to conduction and excitation in nerve”. The Journal of Physiology. 117 (4): 500—544. PMC 1392413 . PMID 12991237. doi:10.1113/jphysiol.1952.sp004764.
- ^ MacDonald PE, Rorsman P (фебруар 2006). „Oscillations, intercellular coupling, and insulin secretion in pancreatic beta cells”. PLOS Biol. 4 (2): e49. PMC 1363709 . PMID 16464129. doi:10.1371/journal.pbio.0040049 .

- ^ Barnett, MW; Larkman, PM (јун 2007). „The action potential”. Pract Neurol. 7 (3): 192—7. PMID 17515599. Архивирано из оригинала 08. 07. 2011. г. Приступљено 01. 11. 2018.
- ^ Mader S. S. (2000): Human biology. Human Biology. New York: McGraw-Hill. 2000. ISBN 978-0-07-290584-7.. ISBN 978-0-07-117940-9.
- ^ Hall, J. E.; Guyton, A. C. (2006). Textbook of medical physiology (11th изд.). St. Louis, Missouri: Elsevier Saunders. ISBN 978-0-7216-0240-0.
- ^ Warrell D. A., Cox T. M., Firth J. D. (2010): The Oxford Textbook of Medicine Архивирано 2012-03-21 на сајту Wayback Machine (5th ed.). Oxford University Press
- ^ Brynie, F.H. (2009). Brain Sense: The Science of the Senses and How We Process the World Around Us. American Management Association.
- ^ Brandt, Thomas (1999). „Vestibular cortex: its locations, functions, and disorders.”. Vertigo. Springer. стр. 219-231. ISBN 9783540199342.
- ^ Duque-Parra JE (септембар 2004). „Perspective on the vestibular cortex throughout history”. Anat Rec B New Anat. 280 (1): 15—9. PMID 15382110. doi:10.1002/ar.b.20031.
- ^ Philbeck JW, Behrmann M, Biega T, Levy L (2006). „Asymmetrical perception of body rotation after unilateral injury to human vestibular cortex”. Neuropsychologia. 44 (10): 1878—90. PMID 16581095. S2CID 58894. doi:10.1016/j.neuropsychologia.2006.02.004.
- ^ Dieterich M, Bense S, Stephan T, Brandt T, Schwaiger M, Bartenstein P (април 2005). „Medial vestibular nucleus lesions in Wallenberg's syndrome cause decreased activity of the contralateral vestibular cortex”. Ann. N. Y. Acad. Sci. 1039 (1): 368—83. Bibcode:2005NYASA1039..368D. PMID 15826990. S2CID 23990811. doi:10.1196/annals.1325.035.
- ^ Kahane P, Hoffmann D, Minotti L, Berthoz A (новембар 2003). „Reappraisal of the human vestibular cortex by cortical electrical stimulation study”. Ann. Neurol. 54 (5): 615—24. PMID 14595651. S2CID 33077726. doi:10.1002/ana.10726.
- ^ Singh, Arun Kumar (септембар 1991). The Comprehensive History of Psychology. Motilal Banarsidass Publ. стр. 66. ISBN 978-81-208-0804-1. Приступљено 8. 4. 2011.
- ^ Dickinson, John (1976). Proprioceptive control of human movement. Princeton Book Co. стр. 4. Приступљено 8. 4. 2011.
- ^ Todd, Robert Bentley (1847). The Cyclopaedia of Anatomy and Physiology Vol. 4. London: Longmans. стр. 585—723.
- ^ McKeeff, T. J.; Tong, F. (2007). „The timing of perceptual decisions for ambiguous face stimuli in the human ventral visual cortex. [Article]”. Cerebral Cortex. 17 (3): 669—678. PMID 16648454. doi:10.1093/cercor/bhk015.
- ^ Hickey, C.; Chelazzi, L.; Theeuwes, J. (2010). „Reward Changes Salience in Human Vision via the Anterior Cingulate. [Article]”. Journal of Neuroscience. 30 (33): 11096—11103. PMID 20720117. doi:10.1523/jneurosci.1026-10.2010.
- ^ Downar, J.; Crawley, A. P.; Mikulis, D. J.; Dav (2000). „multimodal cortical network for the detection of changes in the sensory environment”. Nature Neuroscience. 3 (3): 277—283. PMID 10700261. S2CID 8807081. doi:10.1038/72991.
- ^ Fox, M. D.; Snyder, A. Z.; Barch, D. M.; Gusnard, D. A.; Raichle, M. E. (2005). „Transient BOLD responses at block transitions”. NeuroImage. 28 (4): 956—966. PMID 16043368. S2CID 8552739. doi:10.1016/j.neuroimage.2005.06.025.
- ^ Shulman, G. I.; Tansy, A. P.; Kincade, M.; Petersen, S. E.; McAvoy, M. P.; Corbetta, M. (2002). „Reactivation of Networks Involved in Preparatory States”. Cerebral Cortex. 12 (6): 590—600. PMID 12003859. doi:10.1093/cercor/12.6.590.
- ^ Starr, Philip A.; Barbaro, Nicholas M.; Larson, Paul S. (30. 11. 2008). Neurosurgical Operative Atlas: Functional Neurosurgery. Thieme. стр. 16, 26. ISBN 9781588903990.
- ^ „Temporal Lobe”. Langbrain. Rice University. Приступљено 2. 1. 2011.
- ^ Smith; Kosslyn (2007). Cognitive Psychology: Mind and Brain. New Jersey: Prentice Hall. стр. 21, 194–199, 349. ISBN 9780131825086.
- ^ Cf. Pickles, James O. (2012). An Introduction to the Physiology of Hearing (4th ed.). Bingley, UK: Emerald Group Publishing Limited, pp. 215–217.
- ^ Nakai, Y; Jeong, JW; Brown, EC; Rothermel, R; Kojima, K; Kambara, T; Shah, A; Mittal, S; Sood, S; Asano, E (2017). „Three- and four-dimensional mapping of speech and language in patients with epilepsy”. Brain. 140 (5): 1351—1370. PMC 5405238 . PMID 28334963. doi:10.1093/brain/awx051.
- ^ Moerel, Michelle; De Martino, Federico; Formisano, Elia (29. 7. 2014). „An anatomical and functional topography of human auditory cortical areas”. Frontiers in Neuroscience. 8: 225. PMC 4114190 . PMID 25120426. doi:10.3389/fnins.2014.00225 .
- ^ Rauschecker, Josef P; Scott, Sophie K (26. 5. 2009). „Maps and streams in the auditory cortex: nonhuman primates illuminate human speech processing”. Nature Neuroscience. 12 (6): 718—724. PMC 2846110 . PMID 19471271. doi:10.1038/nn.2331.
- ^ Lush, Mark E.; Piotrowski, Tatjana (2013). „Sensory hair cell regeneration in the zebrafish lateral line”. Developmental Dynamics. 243 (10): 1187—1202. PMC 4177345 . PMID 25045019. doi:10.1002/dvdy.24167.
- ^ Cotanche, Douglas A. (1994). „Hair cell regeneration in the bird cochlea following noise damage or ototoxic drug damage”. Anatomy and Embryology. 189 (1): 1—18. PMID 8192233. S2CID 25619337. doi:10.1007/bf00193125.
- ^ „Sylvius Neuroanatomical Reference”. Архивирано из оригинала 12. 6. 2011. г. Приступљено 20. 11. 2009.
- ^ Zelano C, Montag J, Khan R, Sobel N (2009). „A specialized odor memory buffer in primary olfactory cortex”. PLOS ONE. 4 (3): e4965. Bibcode:2009PLoSO...4.4965Z. PMC 2654926 . PMID 19305509. doi:10.1371/journal.pone.0004965 .
- ^ Johnson DM, Illig KR, Behan M, Haberly LB (септембар 2000). „New features of connectivity in piriform cortex visualized by intracellular injection of pyramidal cells suggest that "primary" olfactory cortex functions like "association" cortex in other sensory systems”. J. Neurosci. 20 (18): 6974—82. PMC 6772836 . PMID 10995842. doi:10.1523/JNEUROSCI.20-18-06974.2000.
- ^ Vermeulen, A; Rospars, J. P. (1998). „Dendritic integration in olfactory sensory neurons: A steady-state analysis of how the neuron structure and neuron environment influence the coding of odor intensity”. Journal of Computational Neuroscience. 5 (3): 243—66. PMID 9663551. S2CID 19598225. doi:10.1023/a:1008826827728.
- ^ Schacter, Daniel L. (2014). Psychology. стр. 167. ISBN 978-1-4641-0603-3.
- ^ Cunningham, A.M.; Manis, P.B.; Reed, R.R.; Ronnett, G.V. (1999). „Olfactory receptor neurons exist as distinct subclasses of immature and mature cells in primary culture”. Neuroscience. 93 (4): 1301—12. PMID 10501454. S2CID 23634746. doi:10.1016/s0306-4522(99)00193-1.
- ^ Touhara, Kazushige (2009). „Insect Olfactory Receptor Complex Functions as a Ligand-gated Ionotropic Channel”. Annals of the New York Academy of Sciences. 1170 (1): 177—80. Bibcode:2009NYASA1170..177T. PMID 19686133. S2CID 6336906. doi:10.1111/j.1749-6632.2009.03935.x.
- ^ Bieri, S.; Monastyrskaia, K; Schilling, B (2004). „Olfactory Receptor Neuron Profiling using Sandalwood Odorants”. Chemical Senses. 29 (6): 483—7. PMID 15269120. doi:10.1093/chemse/bjh050.
- ^ Fan, Jinhong; Ngai, John (2001). „Onset of Odorant Receptor Gene Expression during Olfactory Sensory Neuron Regeneration”. Developmental Biology. 229 (1): 119—27. PMID 11133158. doi:10.1006/dbio.2000.9972.
- ^ Hamilton, K.A.; Heinbockel, T.; Ennis, M.; Szabó, G.; Erdélyi, F.; Hayar, A. (2005). „Properties of external plexiform layer interneurons in mouse olfactory bulb slices”. Neuroscience. 133 (3): 819—829. ISSN 0306-4522. PMC 2383877 . PMID 15896912. doi:10.1016/j.neuroscience.2005.03.008.
- ^ Purves, Dale et al. 2008. Neuroscience. Second Edition. Sinauer Associates Inc. Sunderland, MA.
- ^ Small DM, Green BG (2012). „A Proposed Model of a Flavor Modality”. Ур.: Murray MM, Wallace MT. The Neural Bases of Multisensory Processes. Frontiers in Neuroscience. CRC Press/Taylor & Francis. ISBN 9781439812174. PMID 22593893.
Литература
- „8”. Ganong's Review of Medical Physiology (23rd изд.). TATA McGraw-Hill Lange. 2010. стр. 150. ISBN 978-0-07-067722-7.
- „8”. Ganong's Review of Medical Physiology (23rd изд.). TATA McGraw-Hill Lange. стр. 150. ISBN 978-0-07-067722-7.
- Wiltschko, F.R. & Wiltschko, W. (2012). „Chapter 8 - Magnetoreception”. Ур.: Carlos López-Larrea. Sensing in Nature. Springer. doi:10.1007/978-1-4614-1704-0.
Спољашње везе
- Године 2004. Нобелова награда за Физиологију и медицину (објављена 4. октобра 2004) је додељена Ричарду Акселу и Линди Бак за њихов рад на објашњавању чула мириса. Тај рад је првобитно објављен као заједнички чланак у часопису 1991. године, у коме је описана веома велика фамилија од око хиљаду гена за рецепторе мириса и како су ти рецептори повезани са мозгом.
- Answers to several questions related to senses and human feeling from curious kids
- The Physiology of the Senses tutorial 12 animated chapters on vision, hearing, touch, balance and memory.