Dopamina
Número CAS:51-61-6.
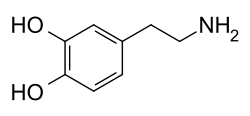
Nome IUPAC: 4-(2-aminoetil)benceno-1,2-diol.
Outros nomes: 2-(3,4-dihidroxifenil)etilamina;
3,4-dihidroxifenetilamina;
3-hidroxitiramina; DA; Intropina; Revivan; Oxitiramina.
A dopamina [1][2][3] (C6H3(OH)2-CH2-CH2-NH2) é un neurotransmisor e unha hormona producida en moitas especies animais tanto vertebrados coma invertebrados incluídos os humanos. Segundo a súa estrutura química, a dopamina é unha catecolamina do grupo das feniletilaminas.
No sistema nervioso, a dopamina cumpre funcións de neurotransmisor, activando os cinco tipos de receptores de dopamina: D1 (relacionado cun efecto activador), D2 (relacionado cun efecto inhibidor), D3, D4 e D5, e as súas variantes. A dopamina prodúcese en moitas partes do sistema nervioso, especialmente na substancia negra. A dopamina é tamén unha neurohormona liberada polo hipotálamo. A súa función principal alí é inhibir a liberación de prolactina no lóbulo anterior da hipófise (adenohipófise).
Como fármaco, actúa como simpaticomimético (emulando a acción do sistema nervioso simpático) promovendo o incremento da frecuencia cardíaca e a presión arterial, e pode producir efectos deleterios como taquicardia ou hipertensión arterial. Porén, debido a que a dopamina non pode atravesar a barreira hematoencefálica, a súa administración como fármaco non afecta directamente ao sistema nervioso central.
A diminución na cantidade de dopamina no cerebro en pacientes con enfermidades como a enfermidade de Parkinson e a distonía en resposta á dopa, L-dopa (Levodopa), que é o precursor da dopamina, pode deberse a que esta última cruza a barreira hematoencefálica. Na enfermidade de Parkinson a destrución das neuronas dopaminérxicas da substancia negra que proxectan cara aos ganglios basais comporta danos tisulares que terminan producindo a perda do control dos movementos a cargo do sistema nervioso.
Historia
A dopamina foi sintetizada artificialmente por vez primeira en 1910 por George Barger e James Ewens nos Laboratorios Wellcome de Londres, Inglaterra. Chamárona dopamina porque é unha monoamina, e o seu precursor sintético é a L-dopa ou 3,4-dihidroxifenilalanina. En 1952, Arvid Carlsson e Nils-Åke Hillarp, do Laboratorio de Farmacoloxía Química do Instituto Nacional do Corazón de Suecia, puxeron de manifesto o seu importante papel como neurotransmisor. Este e outros traballos sobre a transdución de sinais no sistema nervioso valéronlle a Carlsson a concesión do Premio Nobel de Medicina e Fisioloxía en 2000.[4][5]
Bioquímica

Nome e familia
A dopamina ten a fórmula química C6H3(OH)2-CH2-CH2-NH2. O seu nome químico é "4-(2-aminoetil)benceno-1,2-diol" e a súa abreviatura é “DA”. É unha catecolamina do tipo das feniletilaminas. A dopamina é un precursor doutras catecolaminas como a noradrenalina (norepinefrina), e a adrenalina (epinefrina).
Biosíntese
A dopamina biosintetízase no corpo (principalmente no tecido de orixe nerviosa da medula suprarrenal) primeiro pola hidroxilación do aminoácido L-tirosina a L-dopa por medio do encima tirosina 3-monooxixenase, tamén coñecida como tirosina hidroxilase, e despois pola descarboxilación da L-dopa por medio do encima dopa-descarboxilase.[6] Nalgunhas neuronas, a dopamina é transformada en noradrenalina pola dopamina beta-hidroxilase.
Nas neuronas, despois da síntese a dopamina almacénase en vesículas, que se liberan na sinapse en resposta a un impulso eléctrico presináptico.
Síntese

Como fármaco medicinal sintetízase nos laboratorios por desmetilación da 2-(3,4-dimetoxifenil)etilamina (á esquerda) utilizando bromuro de hidróxeno (HBr), rendendo dopamina (á dereita).[7][8]
Inactivación e degradación
A dopamina e inactivada pola recaptación por medio do transportador de dopamina, logo é cortada encimaticamente pola catecol-O-metil transferase (COMT) e a monoamino oxidase (MAO). A dopamina que non é cortada polos encimas é reempaquetada en vesículas para a súa reutilización.
A dopamina tamén pode difundir por difusión simple na sinapse e regular a presión sanguínea.
Funcións no sistema nervioso
A dopamina ten moitas funcións no cerebro, incluíndo papeis importantes no comportamento e a cognición, a actividade motora, a motivación e a recompensa, a regulación da produción de leite, o sono, o estado de ánimo, a atención, e a aprendizaxe.
As neuronas dopaminérxicas (é dicir, as neuronas que utilizan como neurotransmisor primario a dopamina) están presentes maioritariamente na área tegmental ventral do cerebro-medio, a parte compacta da substancia negra, e o núcleo arcuato do hipotálamo.
Segundo unha hipótese a dopamina transmite os erros na predición dunha recompensa, aínda que isto foi cuestionado.[9] De acordo con esta hipótese, obsérvanse respostas fásicas das neuronas dopaminérxicas cando se presenta unha recompensa inesperada. Estas respostas poden establecer un estímulo condicionado despois de que se asocie repetidamente coa recompensa. As neuronas dopaminérxicas son deprimidas cando se omite a recompensa esperada. Así as neuronas dopaminérxicas parecen codificar a predición de erro das recompensas. Na natureza, aprendemos a repetir os comportamentos que levan a maximizar as recompensas. A dopamina crese que proporciona un sinal instrutivo ás partes do cerebro responsables de adquirirmos un novo comportamento.
En insectos, existe un sistema de recompensa similar, que utiliza a octopamina, unha substancia similar á dopamina.[10]
Anatomía
As neuronas dopaminérxicas forman un sistema neurotransmisor que se orixina na parte compacta da substancia negra, a área tegmental ventral e o hipotálamo. Os seus axóns proxéctanse ata varias áreas do cerebro seguindo as seguintes vías principais:
- Vía mesocortical
- Vía mesolímbica
- Vía nigroestriatal
- Vía tuberoinfundibular
Esta innervación explica moitos dos efectos producidos pola activación deste sistema dopaminérxico. Por exemplo, a vía mesolímbica conecta a área tegmental ventral e o núcleo accumbens, ambos os dous son centrais para o sistema de recompensa cerebral.[11]
Aínda que a distinción entre estas vías é moi usada, hai tamén un certo solapamento nas áreas ás que chegan cada unha destes grupos de neuronas.[12]
Efectos celulares
Actividade tónica e fásica
Os niveis de dopamina extracelular están modulados por dous mecanismos: transmisión de dopamina tónica e fásica. A transmisión de dopamina tónica ocorre cando se liberan pequenas cantidades de dopamina independentemente da actividade neuronal, e está regulada pola actividade doutras neuronas e recaptación de neurotransmisores.[13] A liberación de dopamina fásica orixínase pola actividade das propias células que conteñen dopamina. Esta actividade está caracterizada por unha actividade de ritmo irregular de picos únicos, e rápidos pulsos de tipicamente 2-6 picos en rápida sucesión.[14][15] Os pulsos concentrados de actividade causan un grande incremento dos niveis de dopamina extracelular comparados cos que se esperaría do mesmo número de picos distribuídos nun longo período de tempo.[16]
Inhibición da recaptación e liberación sináptica
A cocaína e as anfetaminas inhiben a recaptación da dopamina; porén, inflúen sobre distintos mecanismos de acción. A cocaína é un bloqueador do transportador de dopamina e de noradrenalina que inhibe competitivamente a captación de dopamina e incrementa o tempo de actuación da dopamina e aumenta a sobreabundancia de dopamina (un incremento de ata o 150%). Igual ca a cocaína, as anfetaminas aumentan a concentración de dopamina no espazo sináptico, pero por un mecanismo diferente. As anfetaminas e as metanfetaminas son similares en estrutura á dopamina, e poden entrar na neurona presináptica, forzando ás moléculas de dopamina a saír das súas vesículas sinápticas de almacenamento e pasar ao espazo sináptico facendo que os transportadores de dopamina funcionen en sentido inverso.
Movemento
Por medio dos receptores de dopamina D1, D2, D3, D4 e D5, a dopamina reduce a influencia da vía indirecta, e incrementa as accións da vía directa implicando aos ganglios basais. A biosíntese insuficiente de dopamina nas neuronas dopaminérxicas poden causar a enfermidade de Parkinson, na cal unha persoa perde a habilidade para executar movementos finos e controlados. A activación fásica dopaminérxica parece ser crucial para unha duradeira codificación interna de habilidades motoras (Beck, 2005).
Cognición e córtex frontal
Nos lóbulos frontais, a dopamina controla o fluxo de información desde outras áreas do cerebro. Os trastornos de dopamina nesta rexión do cerebro poden causar un declive nas funcións neurocognitivas, especialmente a memoria, atención, e resolución de problemas. Os niveis reducidos de dopamina no córtex prefrontal pénsase que contribúen ao trastorno por déficit de atención con hiperactividade. Polo contrario, a medicación anti-psicótica actúa como antagonista da dopamina e utilízase no tratamento dos síntomas positivos na esquizofrenia.
Regulación da secreción de prolactina
A dopamina é o principal regulador neuroendócrino da secreción de prolactina pola adenohipófise ou pituitaria anterior. A dopamina producida polas neuronas no núcleo arcuato do hipotálamo é segregada aos vasos sanguíneos hipotálamo-hipofisiarios na eminencia media. As células lactotropas que producen prolactina, en ausencia de dopamina, segregan prolactina continuamente; a dopamina inhibe a súa secreción. Así, no contexto da regulación da secreción de prolactina, a dopamina é ocasionalmente chamada Factor Inhibidor de Prolactina (PIH), ou prolactostatina. A prolactina tamén parece inhibir a liberación de dopamina, como un efecto posterior ao orgasmo, e é principalmente responsable do Período Refractario.
Motivación e pracer
Reforzo
A dopamina é xeralmente asociada co sistema do pracer do cerebro, que produce os sentimentos de gozo e reforzo para motivar a unha persoa a realizar certas actividades. A dopamina que actúa no núcleo accumbens, a amígdala, a área septal lateral, o núcleo olfactorio anterior, o tubérculo olfactorio e o neocórtex libérase polas proxeccións procedentes da área tegmental ventral, e participa na recompensa de experiencias como a alimentación, o sexo,[17][18] algunhas drogas, e os estímulos neutrais que se poden asociar con estes. Esta teoría é frecuentemente discutida en relación con drogas tales como a cocaína, nicotina, e anfetaminas, as cales parecen causar directa ou indirectamente un incremento de dopamina nestas áreas, e en relación coas teorías neurobiolóxicas da adicción química, argumentando que esas vías dopaminérxicas son alteradas patolóxicamente nas persoas adictas. Porén, segundo estudos recentes existe unha relación na alteración nos niveis de dopamina producidos polo tabaco e unha diminución do risco de contraer párkinson, pero os mecanismos de tal relación aínda non se determinaron.[19][20]
Inhibición da recaptación e expulsión
A cocaína e anfetamina inflúen de distinta maneir sobre os mecanismos neuronais. A cocaína é un bloqueador do transportador da dopamina que inhibe competitivamente a recaptación da dopamina para aumentar o período de vida da dopamina e producir unha sobreabundancia de dopamina (un aumento de ata o 150%).
Igual que a cocaína, as anfetaminas incrementan a concentración de dopamina no espazo [sináptico], pero por medio dun mecanismo distinto. As anfetaminas teñen unha estrutura similar á dopamina e poden, por tanto, penetrar no botón terminal da neurona presináptica utilizando os seus transportadores de dopamina, e tamén difundíndose directamente. Ao entrar na neurona presináptica, as anfetaminas forzan as moléculas de dopamina a saíren da súa vesícula de almacenamento e expúlsanas ao espazo sináptico facendo funcionar á inversa os transportadores de dopamina.
O papel da dopamina na experiencia do pracer foi cuestionado por varios investigadores. Argumentouse que a dopamina está máis asociada ao desexo anticipatorio e a motivación por oposición ao pracer consumatorio real.
A dopamina no é liberada cando recibimos estímulos desagradables ou aversivos, e así motiva cara ao pracer de evitar ou eliminar os estímulos desagradables.
Estudos en animais
O que se sabe sobre a dopamina en canto ao seu papel na motivación, o desexo e o pracer, obtívose de estudos realizados en animais. Nun destes estudos con ratas, diminuíuselle ata nun 99% a dopomina no nucleus accumbens e o neostriatum utilizando 6-hidroxidopamina. -->[21] Con esta gran redución de dopamina, as ratas xa non puuderon alimentarse pola súa propia vontade. Os investigadores alimentáronas de maneira forzada e rexistraron as expresións faciais que indicaban se lles agradaba ou non. Concluíron que a redución de dopamina non diminúe o pracer de consumo, só o desexo de comer. Noutro estudo, ratos coa dopamina incrementada mostraron un maior desexo, pero non un maior gusto polas recompensas doces.[22]
Drogas redutoras da dopamina en seres humanos
En humanos, as drogas que reducen a actividade da dopamina (neurolepticos, por exemplo algúns antipsicóticos) reducen tamén a motivación, e provocan anhedonia (incapacidade para experimentar pracer).[23] Contrariamente aos agonistas de D2/D3 pramipexole e ropinirole teñen propiedades anti-anhedónicas, o que foi estimado utilizando a Escala do Pracer de Snaith-Hamilton.[24] (A Escala do Pracer de Snaith-Hamilton, foi introducida en Inglaterra en 1995 para autoavaliar a anhedonia en pacientes psiquiátricos.)
Transmisión cannabinoide e opioide
Os opioides e cannabinoides, en lugar de modular a dopamina poden modular o pracer dos alimentos e a palatabilidade (sabor).[25] Isto podería explicar por que nos animais o "sabor" da comida é independente da concentración de dopamina no cerebro. Outros praceres, porén, poden estar máis asociados coa dopamina. Un estudo informou que tanto a anticipación como a consumación da conduta sexual (machos) foron interrompidas por receptores antagonistas de DA.[26] A libido pode ser incrementada por drogas que afectan á dopamina, pero non por outras que afecten aos péptidos opioides ou doutros neurotransmisores.
Socialización
A sociabilidade está tamén moi ligada á neurotransmisión de dopamina. Unha baixa captabilidade de dopamina no D2 encóntrase con frecuencia en persoas con ansiedade social. Características comúns á esquizofrenia negativa (asociabilidade, apatía, anhedonia) pénsase que están en relación co estado hipodopaminérgico en certas áreas do cerebro. En casos de trastorno bipolar, suxeitos maníacos poden ser hipersociais, igual que tamén poden ser hipersexuais. Isto crese que tamén se produce debido a un incremento de dopamina, debido a que a manía pode ser tratada con antipsicóticos bloqueadores de dopamina.
Saliencia
A dopamina tamén pode ter un papel na chamada saliencia ('perceptibilidade') dos estímulos potencialmente importantes, tales como as fontes de recompensa ou de perigo. Esta hipótese sostén que a dopamina axuda á toma de decisións ao influír na prioridade, ou o nivel de desexo, destes estímulos á persoa en cuestión.
Trastornos do comportamento
O bloqueo dos receptores cerebrais de dopamina aumenta (en vez de diminuír) o consumo de drogas. Dado que o bloqueo de dopamina diminúe o desexo, o aumento no consumo de drogas podería verse non como un desexo químico senón como un profundo desexo psicolóxico de "sentir algo".
O déficit nos niveis de dopamina relacionouse co déficit atencional con hiperactividade (DAH) e os medicamentos estimulantes usados exitosamente para tratar o aumento desmedido nos niveis de neurotransmisores de dopamina levan á diminución dos síntomas.
Inhibición latente e creatividade
A dopamina dos circuítos mesolímbicos incrementa a actividade xeral e a dos centros regulatorios da conduta, diminuíndo a inhibición latente. Estes tres efectos dan como resultado o incremento da creatividade na xeración de ideas. Isto levou ao modelo trifactorial da creatividade que inclúe os lóbulos frontais e temporais e a dopamina mesolímbica. [27]
Hipótese da mente dopaminérxica
A hipótese da mente dopaminérxica trata de explicar as diferenzas entre os humanos modernos e os seus antepasados homínidos centrándose nos cambios no uso da dopamina.[28] A hipótese supón que o incremento dos niveis de dopamina foi parte dunha adaptación fisiolóxica xeral debido ao incremento do consumo de carne hai uns dous millóns de anos por parte do Homo habilis, e despois isto amplificouse polos cambios na dieta e outros factores ambientais e sociais que comezaron hai uns 80.000 anos. Nesta hipótese, a personalidade "alta en dopamina" caracterízase pola elevada intelixencia, un sentido do destino persoal, preocupación cósmica/relixiosa, unha obsesión en conseguir determinadas fins e conquistas, un desapego emocional que en moitos casos leva á crueldade, e unha mentalidade favorable á toma de riscos. Proponse que os altos niveis de dopamina están detrás dun incremento dos trastornos psicolóxicos nas sociedades industriais. De acordo con esta hipótese, unha "sociedade dopaminérxica" está estremadamente orientada a conseguir fins, ten un ritmo acelerado e mesmo características maníacas, "dado que se sabe que a dopamina incrementa os niveis de actividade, acelera os nosos reloxos internos e crea unha preferencia por ambientes novos en vez de sen cambio." [28] Do mesmo modo que os individuos con alta dopamina perden a empatía e mostran un estilo de comportamento máis masculino, as sociedades dopaminérxicas están "tipificadas por máis conquistas, competición, e agresividade."[28] Aínda que a hipótese está apoiada por evidencias de comportamento e algunhas evidencias anatómicas indirectas (por exemplo, aumento nos humanos do striatum rico en dopamina)[29], aínda non hai unha evidencia directa de que os niveis de dopamina sexan marcadamente máis altos nos humanos ca nos grandes simios.[30] Porén, recentes descubrimentos sobre asentamentos á beira do mar dos primeiros homes poden ser unha evidencia de cambios na dieta que apoiarían esta hipótese.[31]
Relación coa psicose
A dopamina anormalmente alta asóciase coa psicose e esquizofrenia [32], aínda que algúns estudos contradín isto.[33] As neuronas de dopamina na vía mesolímbica están particularmente asociadas con estes síntomas. As probas proceden parcialmente do descubrimento dunha clase de drogas chamadas fenotiacinas (que bloquean os receptores de dopamina D2) que poden reducir os síntomas psicóticos, e parcialmente do descubrimento de que drogas como a anfetamina e cocaína (que son coñecidas por incrementar de maneira importante os receptores de dopamina) poden causar psicose.[34] Por esta razón, a maioría dos modernos fármacos antipsicóticos, por exemplo, Risperidona, están deseñados para bloquear a función da dopamina en diversos graos.
Uso terapéutico
- Artigo principal: Levodopa.
A levodopa é un precursor da dopamina usado de varias maneiras no tratamento da enfermidade de Parkinson. Coadminístrase normalmente xunto cun inhibidor da descarboxilación periférica (DDC, dopa descarboxilase), como a carbidopa ou benserazida. Tamén se usan os inhibidores da ruta metabólica alternativa da dopamina pola catecol-O-metil transferase, entre os que están a entacapona e tolcapona.
Control das funcións renais e cardiovasculares
A dopamina ten tamén efectos fóra do sistema nervioso central cando se admministra por vía intravenosa IV, que son dependentes da dose.
- Induce a natriurese (eliminación de sodio) nos riles a doses baixas, e ten un efecto diurético.[35][36][37][38]
- A doses intermedias ten ademais un efecto positivo inotrópico e cronotrópico por medio do incremento da activación dos receptores β1. A dopamina utilízase en pacientes con shock circulatorio ou insuficiencia cardíaca para incrementar o gasto cardíaco e a presión arterial.[38][39]
- A altas doses [40] causa vasoconstrición, incrementa a resistencia vascular sistémica, e a presión sanguínea ao activar os receptores α1,[38] pero produce constrición dos vasos renais e redución do volume de urina.[40]
Funcións inmunorregulatorias
A dopamina actúa sobre os receptores presentes nas células inmunes, con todos os subtipos de receptores de dopamina presentes nos leucocitos. Hai unha expresión baixa destes receptores no linfocito T e no monocito, unha expresión moderada no neutrófilo e eosinófilo, e unha alta expresión no linfocito B e na célula NK.[41] A innervación simpática do tecido linfoide é dopaminérxica, e increméntase durante o estrés.[42] A dopamina pode tamén afectar ás células inmunitarias do bazo, medula ósea, e sangue.[43] Ademais, a dopamina poden sintetizala e liberala as propias células inmunitarias.[44][45]
Os efectos da dopamina sobre as células inmunitarias dependen do seu estado fisiolóxico. A dopamina activa as células T en repouso, pero inhíbeas cando están activadas. Trastornos como a esquizofrenia e enfermidade de Parkinson, nas cales hai cambios nos receptores de dopamina do cerebro e nas vías de sinalización de dopamina, están tamén asociados coa alteración das funcións inmunes.[46]
A dopamina e a oxidación da froita
As polifenol oxidases (PPOs) son unha familia de encimas responsables da oxidación de froitas frescas e vexetais despois de seren cortados ou mazados. Estes encimas usan O2 para oxidar varios difenois ás súas correspondentes quinonas. O substrato natural para os PPOs na banana é a dopamina. O produto da súa oxidación, a dopamina quinona, oxídase espontaneamente en presenza doutras quinonas. As quinonas entón polimerizan e condensan con aminoácidos para formar pigmentos castaños denominados melaninas. Crese que estas quinonas e melaninas derivadas da dopamina poderían axudar a protexer as froitas e vexetais danados das bacterias e fungos.[47]
Notas
- ↑ ChemSpider Dopamine
- ↑ PubChem compound Dopamine
- ↑ CHEBI
- ↑ Benes, F.M. (1 de enero de 2001). "Carlsson and the discovery of dopamine". Trends in Pharmacological Sciences. Volume 22 (Issue 1): 46–47.
- ↑ Fahn, Stanley (1º de noviembre de 2006). The Movement Disorder Society, ed. "The History of Levodopa as it Pertains to Parkinson’s Disease" (PDF). 10th international congress of parkinson’s disease and movement disorders (Kyoto, Japón). Arquivado dende o orixinal (PDF) o 29 de marzo de 2012. Consultado o 07 de abril de 2012.
- ↑ Zhen Qi, Gary W. Miller, Eberhard O. Voit (2008). Computational Systems Analysis of Dopamine Metabolism (artigo completo dispoñible en inglés). PLoS ONE 3(6): e2444. doi:10.1371/journal.pone.0002444 Último acceso 26 de xuño de 2010.
- ↑ J. S. Bayeler, Ann. Chem., 513, 196 (1934).
- ↑ G. Hahn, K. Stiehl, Chem. Ber., 69, 2640 (1936).
- ↑ Peter Redgrave, Kevin Gurney (2006). "The short-latency dopamine signal: a role in discovering novel actions?". Nature Reviews Neuroscience 7 (12): 967–975. PMID 17115078. doi:10.1038/nrn2022.
- ↑ Barron AB, Maleszka R, Vander Meer RK, Robinson GE (2007). "Octopamine modulates honey bee dance behavior". Proc. Natl. Acad. Sci. U.S.A. 104 (5): 1703–7. PMID 17237217. doi:10.1073/pnas.0610506104.
- ↑ "Schultz, Cambridge university, UK". Arquivado dende o orixinal o 11 de abril de 2012. Consultado o 07 de abril de 2012.
- ↑ Bjorklund A, Dunnett SB (2007). "Dopamine neuron systems in the brain: an update". Trends in Neurosciences 30 (5): 194–202. PMID 17408759. doi:10.1016/j.tins.2007.03.006.
- ↑ Grace AA, (1991). "Phasic versus tonic dopamine release and the modulation of dopamine system responsivity: A hypothesis for the eitiology of schizophrenia". Neuroscience 41 (1): 1–24. PMID 1676137. doi:10.1016/0306-4522(91)90196-U.
- ↑ Grace AA, Bunney BS (1984). "The control of firing pattern in nigral dopamine neurons: single spike firing" (pdf). Journal of Neuroscience 4 (11): 2866–2876. PMID 6150070.
- ↑ Grace AA, Bunney BS (1984). "The control of firing pattern in nigral dopamine neurons: burst firing" (pdf). Journal of Neuroscience 4 (11): 28677–2890. PMID 6150071.
- ↑ Gonon FG (1988). "Nonlinear relationship between impulse flow and dopamine released by rat midbrain dopaminergic neurons as studied by in vivo electrochemistry". Neuroscience 24 (1): 19–28. PMID 3368048. doi:10.1016/0306-4522(88)90307-7.
- ↑ Giuliano, F.; Allard J. (2001). "Dopamine and male sexual function". Eur Urol 40: 601–608. PMID 11805404.
- ↑ Giuliano, F.; Allard J. (2001). "Dopamine and sexual function". Int J Impot Res 13 (Suppl 3): S18–S28. PMID 11477488. doi:10.1038/sj.ijir.3900719.
- ↑ Ma, Chaoran; Liu, Yesong; Neumann, Samantha; Gao, Xiang (2017-07-02). "Nicotine from cigarette smoking and diet and Parkinson disease: a review". Translational Neurodegeneration 6. ISSN 2047-9158. PMC 5494127. PMID 28680589. doi:10.1186/s40035-017-0090-8.
- ↑ "Parkinson's disease: the protective role of cigarette smoking is based on interactions with certain genes". Inserm - From science to health (en inglés). Arquivado dende o orixinal o 25 de marzo de 2019. Consultado o 25 de marzo de 2019.
- ↑ Berridge K, Robinson T (1998). "What is the role of dopamine in reward: hedonic impact, reward learning, or incentive salience?". Brain Res Brain Res Rev 28 (3): 309–69. PMID 9858756.
- ↑ Peciña S, Cagniard B, Berridge K, Aldridge J, Zhuang X (2003). "Hyperdopaminergic mutant mice have higher "wanting" but not "liking" for sweet rewards.". J Neurosci 23 (28): 9395–402. PMID 14561867.
- ↑ Lambert M, Schimmelmann B, Karow A, Naber D (2003). "Subjective well-being and initial dysphoric reaction under antipsychotic drugs - concepts, measurement and clinical relevance.". Pharmacopsychiatry 36 (Suppl 3): S181–90. PMID 14677077.
- ↑ Lemke M, Brecht H, Koester J, Kraus P, Reichmann H (2005). "Anhedonia, depression, and motor functioning in Parkinson's disease during treatment with pramipexole.". J Neuropsychiatry Clin Neurosci 17 (2): 214–20. PMID 15939976. Arquivado dende o orixinal o 27 de setembro de 2011. Consultado o 07 de abril de 2012.
- ↑ Peciña S, Berridge K (2005). "Hedonic hot spot in nucleus accumbens shell: where do mu-opioids cause increased hedonic impact of sweetness?". J Neurosci 25 (50): 11777–86. PMID 16354936.
- ↑ Pfaus J, Phillips A (1991). "Role of dopamine in anticipatory and consummatory aspects of sexual behavior in the male rat.". Behav Neurosci 105 (5): 727–43. PMID 1840012.
- ↑ Flaherty, A.W, (2005). "Frontotemporal and dopaminergic control of idea generation and creative drive". Journal of Comparative Neurology 493 (1): 147–153. PMID 16254989.
- ↑ 28,0 28,1 28,2 Previc F (2009). The Dopaminergic Mind in Human Evolution and History Cambridge University Press. ISBN 978-0-521-51699-0.
- ↑ Rapoport, S. I. (1990). "Integrated phylogeny of the primate brain, with special reference to humans and their diseases". Brain Research Reviews 15 (3): 267–294. PMID 2289087. doi:10.1016/0165-0173(90)90004-8.
- ↑ Raghanti, M. A.; Stimpson, C. D.; Marcinkiewicz, J. L.; Erwin, J. M.; Hof, P. R.; Sherwood, C. C. (2008a). "Cortical dopaminergic innervation among humans, chimpanzees, and macaque monkeys: A comparative study". Neuroscience 155 (1): 203–20. PMC 3177596. PMID 18562124. doi:10.1016/j.neuroscience.2008.05.008.
- ↑ "Copia arquivada". Arquivado dende o orixinal o 26 de decembro de 2010. Consultado o 06 de maio de 2012.
- ↑ "Disruption of gene interaction linked to schizophrenia". St. Jude Children's Research Hospital. Consultado o 06-07-2006.
- ↑ Maas, J.W.; Bowden CL, Miller AL, Javors MA, Funderburg LG, Berman N, Weintraub ST. (1997). "Schizophrenia, psychosis, and cerebral spinal fluid homovanillic acid concentrations.". Schizophrenia Bulletin. 23 (1): 147–154. PMID 9050120.
- ↑ Lieberman, J.A.; JM Kane, J. Alvir (1997). "Provocative tests with psychostimulant drugs in schizophrenia.". Psychopharmacology (Berl). 91 (4): 415–433. PMID 2884687. Consultado o 2007-10-16.
- ↑ Gildea, John J (2009). "Dopamine and angiotensin as renal counterregulatory systems controlling sodium balance". Current Opinion in Nephrology and Hypertension 18 (1): 28–32. PMC 2847451. PMID 19077686. doi:10.1097/MNH.0b013e32831a9e0b.
- ↑ Cinelli, A. R.; Efendiev, R.; Pedemonte, C. H. (2008). "Trafficking of Na-K-ATPase and dopamine receptor molecules induced by changes in intracellular sodium concentration of renal epithelial cells". AJP: Renal Physiology 295 (4): F1117–25. PMC 2576148. PMID 18701625. doi:10.1152/ajprenal.90317.2008.
- ↑ "Renal Vasodilatory Action of Dopamine in Patients With Heart Failure: Magnitude of Effect and Site of Action". Circulation. 2008;117:200-205. Arquivado dende o orixinal o 13 de febreiro de 2010. Consultado o 2009-04-20.
- ↑ 38,0 38,1 38,2 Shen, Howard (2008). Illustrated Pharmacology Memory Cards: PharMnemonics. Minireview. p. 8. ISBN 1-59541-101-1.
- ↑ "Dopamine and Dextrose". Drugs.com. Consultado o 2009-04-20.
- ↑ 40,0 40,1 Jean Bryant Bronwen, Kathleen Mary Knights (15 November 2009). Elsevier Australia, ed. Pharmacology for Health Professionals (2nd ed.). p. 192. ISBN 978-0-7295-3929-6. Consultado o 9 June 2011.
- ↑ McKenna, F; McLaughlin, PJ; Lewis, BJ; Sibbring, GC; Cummerson, JA; Bowen-Jones, D; Moots, RJ. (2002). "Dopamine receptor expression on human T- and B-lymphocytes, monocytes, neutrophils, eosinophils and NK cells: a flow cytometric study". J Neuroimmunol 132 (1–2): 34–40. PMID 12417431. doi:10.1016/S0165-5728(02)00280-1.
- ↑ Mignini, F; Streccioni, V; Amenta, F (2003). "Autonomic innervation of immune organs and neuroimmune modulation". Autonomic & autacoid pharmacology 23 (1): 1–25. PMID 14565534. doi:10.1046/j.1474-8673.2003.00280.x.
- ↑ Basu, S; Dasgupta, PS. (2000). "Dopamine, a neurotransmitter, influences the immune system". J Neuroimmunol 102 (2): 113–24. PMID 10636479. doi:10.1016/S0165-5728(99)00176-9.
- ↑ Bergquist, J; Tarkowski, A; Ekman, R; Ewing, A. (1994). "Discovery of endogenous catecholamines in lymphocytes and evidence for catecholamine regulation of lymphocyte function via an autocrine loop". Proc Natl Acad Sci U S A 91 (26): 12912–6. PMC 45550. PMID 7809145. doi:10.1073/pnas.91.26.12912.
- ↑ Cosentino, M; Fietta, AM; Ferrari, M; Rasini, E; Bombelli, R; Carcano, E; Saporiti, F; Meloni, F; Marino, F (2007). "Human CD4+CD25+ regulatory T cells selectively express tyrosine hydroxylase and contain endogenous catecholamines subserving an autocrine/paracrine inhibitory functional loop". Blood 109 (2): 632–42. PMID 16985181. doi:10.1182/blood-2006-01-028423. Arquivado dende o orixinal o 29-05-2010. Consultado o 06-05-2012.
- ↑ Sarkar, C; Basu, B; Chakroborty, D; Dasgupta, PS; Basu, S (2010). "The immunoregulatory role of dopamine: an update". Brain, behavior, and immunity 24 (4): 525–8. PMC 2856781. PMID 19896530. doi:10.1016/j.bbi.2009.10.015.
- ↑ Mayer, AM (2006). "Polyphenol oxidases in plants and fungi: Going places? A review". Phytochemistry 67: 2318–2331. PMID 16973188.