Plantenfysiologie

Plantenfysiologie is de biologische wetenschap die de levensverrichtingen van planten bestudeert. Het onderzoek naar de levensfuncties van planten omvat aspecten zoals het metabolisme, organisatie, de waterhuishouding en transport, de groei en ontwikkeling, de reactie op het milieu en de hormonale regeling.[1] Veel principes van de plantenfysiologie komen voort uit de scheikunde, fysische chemie en natuurkunde. Kennis van plantenfysiologie is van groot belang vanwege de toepassingen in andere disciplines, zoals landbouw, bosbouw en horticultuur.[2]
Een plant gebruikt zonlicht, koolstofdioxide uit de lucht, en water en mineralen uit de bodem om te groeien en in leven te blijven. Tijdens de groei vormt de plant verschillende gespecialiseerde cellen, weefsels en organen die ervoor zorgen dat de plant duurzaam in stand kan blijven en zich kan voortplanten. De precieze mechanismen waarmee een plant deze processen uitvoert zijn nog niet geheel ontrafeld. Het is echter duidelijk geworden dat de fysiologie afhankelijk is van een fijnzinnige wisselwerking tussen een aantal interne en externe factoren.
Planten onderscheiden zich van de dieren en schimmels door hun unieke biochemische en cellulaire organisatie. Door middel van fotosynthese, het moleculaire proces waarmee energie uit licht wordt gevangen en gebruikt wordt voor de vorming van suikers, zijn planten in staat zichzelf te voorzien van energie. Planten produceren een breed scala aan natuurproducten die niet voorkomen in andere organismen, waaronder pigmenten voor fotosynthese, toxines tegen herbivoren en pathogenen, en hormonen die groei reguleren.
Het leven van planten
Karakteristieken van planten

Planten vormen een rijk van meercellige eukaryoten. Planten onderscheiden zich van de schimmels en de dieren door hun vermogen tot fotosynthese: planten halen het grootste deel van hun energie uit zonlicht en abiotische componenten. In chloroplasten, de organellen waarin fotosynthese plaatsvindt, komen chlorofyllen voor die de plant zijn groene uiterlijk geven. Planten vertonen een generatiewisseling in hun levenscyclus: de meeste hebben een opvallende diploïde (sporofyt) en minder opvallende haploïde (gametofyt) kernfase.
Er zijn ongeveer 320.000 plantensoorten beschreven, waarvan veruit de meerderheid, zo'n 260-290 duizend, zaden vormen.[3] Groene planten produceren een aanzienlijk deel van de zuurstof in de atmosfeer en staan aan de basis van de meeste ecosystemen.[4] Planten die graan, fruit en groenten produceren, vormen ook het basisvoedsel voor de mens en zijn om die reden al duizenden jaren geleden gedomesticeerd. Gedomesticeerde dieren, met name de landbouwhuishouddieren, worden gevoerd met planten en plantaardige producten.
Sessiliteit
Planten zijn sessiele organismen, wat inhoudt dat ze hun leven lang verankerd zijn in de grond of een ander substraat, beweeglijkheid (motiliteit) komt niet voor in het plantenrijk. Gedurende de evolutie hebben planten zich op verschillende manieren aangepast aan hun sessiele levenswijze, onder andere door hun onbepaalde groei, hun modulaire bouw, het vaak sterke vermogen tot ongeslachtelijke voortplanting, hun vermogen tot maximale opvang van zonlicht.
Planten hebben veelal een onbepaalde groei gedurende hun levenscyclus: ze zijn in staat om onophoudelijk naar het licht te groeien, vaak tegen de zwaartekracht in. Ook wortels kunnen zich ondergronds voortdurend uitbreiden en kunnen op deze manier steeds nieuwe plaatsen bereiken. Planten kunnen zelfs hun voedsel opzoeken op nieuwe plaatsen, bijvoorbeeld met rizomen en uitlopers.
Planten zijn modulair gebouwd met een klein aantal basisonderdelen: wortel, stengel en blad. Steeds worden nieuwe organen gevormd en sterven andere af zonder nadelige gevolgen voor de plant. De organen van planten hebben een grote milieuafhankelijke plasticiteit, waardoor ze zich binnen hun leven kunnen aanpassen aan de omgeving.
Een aantal plantensoorten heeft een goed ontwikkeld vermogen om zich ongeslachtelijk voort te planten. Bij slechte voorziening met nutriënten kunnen planten hun snelheid van ongeslachtelijke voortplanting verhogen.
De bladstand is zodanig dat de bladeren weinig onderlinge beschaduwing veroorzaken, zodat er maximaal licht gevangen kan worden. Ook de stand van de bladschijf is gewoonlijk zodanig dat veel licht wordt opgevangen, of is de anatomische bouw van de bladeren afhankelijk van de positie aan de plant en de mate van beschaduwing.
Voortplanting en vestiging
Een sessiel organisme kan niet op zoek gaan naar een partner voor de overdracht van gameten of de nakomelingen gemakkelijk verspreiden. Planten lossen het probleem van de gametenoverdracht op door te vertrouwen op verschillende vectoren voor de bestuiving, zoals wind, water, insecten of andere dieren. Mechanismen voor de verspreiding van vruchten en zaden helpen de nakomelingen zich te verspreiden.
Sessiliteit brengt eveneens met zich mee dat planten niet in staat zijn om zich te verplaatsen naar een gunstiger locatie.[a] De (meestal toevallige) plaats van kieming is bepalend voor het verdere verloop van het leven. Veel planten produceren dan ook veel nakomelingen, waarvan er maar weinig overleven. Kieming is over het algemeen alleen mogelijk onder lichte omstandigheden, dus bij zaden die niet te diep onder het grondoppervlakte liggen. Er zijn echter ook zaden van bepaalde plantensoorten die voor de kieming juist duisternis nodig hebben, zoals de bolderik.
Grotere zaden kiemen soms pas bij voldoende hoge concentraties van het plantenhormoon etheen, dat door de plant en door micro-organismen wordt geproduceerd. Sommige planten hebben gespecialiseerde mechanismen voor de verspreiding, wat de kans van het zaad zal verhogen om op de juiste plaats terecht te komen. De kleverige bessen van de maretak worden bijvoorbeeld verspreid door vogels.
Plantencellen

Plantencellen zijn opgebouwd uit de protoplast of het cytoplasma (de levende inhoud van de cel), omgeven door een celwand (een betrekkelijk star omhulsel).[5] De protoplast bevat een aantal inwendige compartimenten en organische moleculen die nodig zijn voor alle levensprocessen van de plant. Planten zijn eukaryotische organismen en hebben een complexe inwendige structuur met verschillende typen organellen.
Het cytoplasma van een plantaardige cel bestaat globaal uit het cytosol (de gelachtige grondvloeistof) met daarin ingebed de celkern met het meeste genetische materiaal, een grote vacuole voor wateropslag, plastiden als chloroplasten en zetmeelkorrels, en diverse andere organellen. Binnen de cel, vaak in de vacuole, zijn ook veel organische stoffen als tannines, kleurstoffen en secundaire metabolieten in de vorm van kristallen aanwezig. Samenwerking tussen de verschillende organellen ligt aan de basis van alle cellulaire activiteit. Het cytoskelet, een vezelig netwerk, geeft de plantencel interne stabiliteit.
De cellen van planten zijn aan elkaar bevestigd met een uit pectine (polysachariden) bestaande middenlamel: het is een tussengelegen laag die naburige cellen aan elkaar verbindt. In de celwanden bevinden zich de plasmodesmata (openingen), die zorgen voor een cytoplasmatische brug tussen aangrenzende cellen. Op deze manier loopt de celmembraan ononderbroken van de ene cel naar de volgende. De celwanden worden later op deze middenlamel afgezet, en bevinden zich ook buiten de celmembraan. Tussen de cellen, waar de celwanden door oplossen van de middenlamel van elkaar wijken, bevinden zich de intercellulaire ruimten, die een functie kunnen hebben bij het transport van gassen of van water.
Protoplast
Een levende plantencel wordt altijd omgeven door een plasmamembraan of celmembraan. Deze bestaat uit een dubbele laag van fosfolipiden met diverse tussengevoegde eiwitten. Fosfolipiden zijn moleculen die zowel hydrofiele als hydrofobe gebieden hebben. Ze hebben een ruggengraat van glycerol met een van de hydroxylgroepen gebonden aan een fosfaat en een geladen groep, de andere twee hydroxylgroepen zijn veresterd met vetzuren. Het plasmamembraan beschermt de levende celinhoud en voorkomt dat macromoleculen vrijelijk wegdiffunderen door bepaalde stoffen selectief door te laten.

Plantencellen beschikken over organellen die ook voorkomen in de cellen van dieren en schimmels, de andere eukaryoten. De belangrijkste organellen zijn de celkern, de mitochondriën, de ribosomen, het endoplasmatisch reticulum en het golgicomplex.
De celkern is omgeven door de kernmembraan: een dubbel membraan bestaande uit twee lipide dubbellagen. In de kernmembraan zijn duizenden kernporiën aanwezig, die bij planten uit meer dan honderd eiwitten bestaan. In de celkern zijn de chromosomen of het chromatine opgeslagen: het erfelijk materiaal van de celkern. Ook is er een nucleolus te herkennen: dit is de plaats waar ribosomen gevormd worden. Binnen het kernplasma komt een netwerk van intermediaire filamenten voor: het kernlamina.
De mitochondriën zijn net als de kern en de plastiden ook voorzien van een dubbel membraan. Deze organellen hebben eigen erfelijk matreriaal (resp. chondroom en plastoom). De binnenste membraan van de mitochondriën is geplooid en dus groter dan de buitenmembraan. De binnenste membraan verschilt van de plasmamembraan, doordat zij een hoger eiwitgehalte met unieke fosfolipiden heeft. Mitochondriën vermenigvuldigen zich door binaire deling en hebben eigen DNA, dat cirkelvormig is, en ribosomen van een ander type dan in de cytosol. Mitochondriën zijn de plaats van de celademhaling; er wordt energie (ATP) vrijgemaakt door omzetting van glucose en vetzuren.
Ribosomen zijn de kleine moleculaire complexen die de eiwitsynthese verzorgen. Ze bestaan uit twee delen, een groter en een kleiner onderdeel. Er zijn vrije ribosomen (deze produceren eiwitten voor binnen de cel), en aan het ruw endoplasmatisch reticulum gebonden ribosomen, die dienen voor de aanmaak van eiwitten voor de export.
Het endoplasmatisch reticulum en golgicomplex zijn verantwoordelijk voor de synthese, modificatie en lokalisatie van eiwitten.
Plastiden
Plastiden zijn voor planten en algen kenmerkende organellen. Ze worden omgeven door een dubbel membraan en ze bevatten hun eigen DNA (het plastoom) en eigen ribosomen. Ze zijn semi-autonoom en planten zich binnen de cel voort door binaire deling. Plastiden worden vrijwel altijd met het cytoplasma van de eicel overgeërfd, maar er zijn voorbeelden van overerving van plastiden via de mannelijke lijn. De chemische samenstelling van de membranen van de plastiden verschilt van de celmembraan.
Er zijn verschillende typen plastiden die verschillende functies kunnen hebben, zoals proplastiden, chloroplasten, chromoplasten, amyloplasten, leukoplasten en etioplasten. Plastiden kunnen opnieuw differentiëren en worden omgevormd van de ene vorm in de andere.
Vacuole
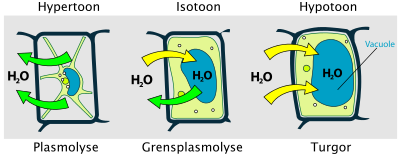
Vacuolen zijn grote, met vocht gevulde blazen met een enkele membraan: de tonoplast of vacuolemembraan. De vacuole maakt 90% of meer uit van het volume van de uitgegroeide cel. Uitzonderingen hierop worden gevormd door embryonale cellen in meristemen. De centrale vacuole bevat water, ionen, organische zuren, suikers, hydrolytische enzymen (zoals ribonucleasen en glycosidasen), en een verscheidenheid van vaak kristallijne, secundaire metabolieten. Vacuolen zijn ook van belang bij het opslaan van afvalstoffen en eiwitten.[b]
De vacuole dient als opslagplaats voor waterstofionen (H+): de tonoplast bevat daarvoor een actieve protonpomp. Hoewel de pH van de cytosol ongeveer bij 7,0 ligt, is de vacuole veel zuurder, met een pH van 2 tot 6. Verder is de vacuole van groot belang bij de lengtegroei van cellen: bij de celstrekking neemt de vacuole sterk toe in grootte maar niet de cytosol die als een dunne laag om de vacuole komt te liggen. De vacuole houdt de turgordruk in plantencellen in stand via de druksterkte van water tegen de stevige celwand.
Celwand

De celwand is een stevige structuur die om de plantencel is gelegen. De celwand bepaalt de vorm van de cel (vergelijkbaar met een uitwendig skelet) en voorkomt dat de celmembraan barst omdat het tegen waterdruk is bestand. Het zorgt voor de ondersteuning van de plant en de mechanische sterkte. De celwand beïnvloedt de snelheid en richting van celgroei, regelt het celvolume, en is uiteindelijk verantwoordelijk voor het architectonisch ontwerp van de plant en voor de besturing van de plantmorfogenese omdat de celwand het voor planten noodzakelijk maakt zich te ontwikkelen door celvorming (en niet door celmigratie).
De celwand is daarnaast een fysieke barrière voor ziekteverwekkers. Kleine moleculen kunnen zich echter door de stippels bewegen. Celwanden hebben naast stevigheid een functie bij de opslag van koolhydraten, vooral in zaden. Fragmenten van de celwand, met name de oligosachariden, kunnen werken als plantenhormonen, bijvoorbeeld bij de productie van etheen, fytoalexine, chitinase en andere enzymen.
De componenten van de celwand kunnen zeer divers zijn:
- De belangrijkste component van celwanden is cellulose of β-1,4-glucaan, bestaande uit lange ketens van glucose. De basale bouwsteen is cellobiose (een disacharide bestaande uit twee glucosemoleculen).
- Hemicellulose is een verzamelnaam voor een groep heteropolymeren, die in tegenstelling tot cellulose, een amorfe structuur aannemen met weinig hechtheid. Twee veel voorkomende soorten zijn xyloglucanen en glucuronarabinoxylanen.
- Pectines zijn ook een diverse groep van polysachariden. Pectines zijn bijzonder rijk aan galacturonzuren. Ze vormen gels, zodat ze vaak gebruikt worden in geleiproducten.
- De eiwitten in de celwand zijn meestal glycoproteïnen (polypeptidehoofdketen met koolhydraatzijketens). De eiwitten kunnen bijzonder rijk zijn aan de aminozuren hydroxyproline, proline en glycine.
- Lignine is een polymeer van fenolen, vooral fenylpropanoïden. Lignines zijn door hun rigiditeit vooral van belang in hout en schors. Ze bieden bestendigheid tegen schimmels en andere ziekteverwekkers.
- Suberine, was en cutine zijn aan de celwand toegevoegde lipiden, die een functie hebben voor de sterkte en de waterdichtheid van de celwand.
De ultrastructuur van de celwand varieert sterk. In de celwand zijn te onderscheiden:
- de middenlamel: de buitenst dunne laag, voornamelijk bestaande uit pectines.
- de primaire celwand: afgezet door de cel tijdens zijn groei. Alle plantencellen hebben een middenlamel en een primaire wand.
- de secundaire celwand: sommige cellen zetten extra lagen af op de primaire wand. Dit gebeurt nadat de groei gestopt is of wanneer de cellen zich beginnen te differentiëren (specialisatie). De secundaire wand is hoofdzakelijk voor versteviging en bestaat voornamelijk uit cellulose en lignine.
- stippels: dunne plekken in de celwand, waardoor de uitwisseling van stoffen tussen cellen gemakkelijker kan plaatsvinden.
Op grond van het type verdikking van de secundaire celwand onderscheidt men collenchym (met onregelmatige, elkaar afwisselende laagjes cellulose en protopectine) en sklerenchym (met gelijkmatig verdikte, sterk verhoute celwanden).
Water en transport
Water

Water is een essentieel gegeven voor alle levende organismen, en de bestudering ervan vormt een grondbeginsel van de plantenfysiologie.[6] De meeste organismen bestaan uit ten minste 70% water; sommige plantendelen, zoals een krop sla, bestaan uit bijna 95% water. Water is van vitaal belang als een oplosmiddel, een stof waarin verbindingen kunnen worden opgelost en getransporteerd worden, maar ook als reactant in stofwisselingsprocessen. Het grote belang van water hangt samen met een aantal bijzondere scheikundige en natuurkundige eigenschappen.
Water is een polaire verbinding, wat betekent dat het gemakkelijk waterstofbruggen vormt. Het watermolecuul is een dipool en heeft een positief geladen (waterstof) en negatief geladen zijde (zuurstof). Door de polariteit van het watermolecuul trekken deze moleculen elkaar sterk aan, wat het ten opzichte van andere stoffen lage smeltpunt, hoge kookpunt en de hoge smeltwarmte en verdampingswarmte verklaart. De polariteit van water maakt ook dat zouten (ionen) en polaire stoffen (zoals suiker) goed in water oplossen.
Fysische eigenschappen
- Bij fysiologische temperaturen is water vloeibaar; water heeft een hoog kookpunt en een hoog smeltpunt in vergelijking met moleculen van vergelijkbare grootte zoals ammonium, koolstofdioxide, waterstofsulfide. Elk van deze stoffen zijn gassen bij kamertemperatuur; water is dat niet.
- Water heeft een hoge verdampingswarmte. Er is relatief veel energie nodig (ca. 44 kJ mol−1) om water om te zetten van een vloeistof naar een gas. Met andere woorden: "water gaat verdamping tegen".
- Water heeft een hoge soortelijke warmte. Het kost veel energie (4,184 J g−1 C−1 bij vloeibaar water, 2,06 J g−1 C−1 bij ijs van 0 °C) om water te verwarmen, omdat het veel energie vereist de waterstofbruggen te breken of te maken. Met andere woorden: "water gaat temperatuurschommelingen tegen". Vetplanten maken gebruik van deze bufferende werking.
- Water heeft een hoge oppervlaktespanning. Het kost veel energie om door de oppervlakte van het water te breken, omdat watermoleculen bij het wateroppervlak veel meer door andere watermoleculen worden aangetrokken (cohesie) dan de moleculen in de lucht. Een wateroppervlak gedraagt zich als een veerkrachtige laag. Dit effect is belangrijk voor het transport van water door de weefsels van een plant.
- Water is lichtdoorlatend, wat van belang is, omdat planten lichtenergie gebruiken voor de syntese van voedingsstoffen. Vanuit een ecologisch perspectief is de doorlaatbaarheid water bepalend voor het voorkomen van de waterplanten.
Chemische eigenschappen
Water beïnvloedt de vorm, de stabiliteit en de eigenschappen van veel biologische moleculen. De meeste ionen (zoals Na+) en macromoleculen (zoals DNA, eiwitten en polysachariden) zijn normaal gesproken gehydrateerd. Dit betekent dat watermoleculen door middel van waterstofbruggen aan deze moleculen gebonden zijn, en sommige gevallen (bijvoorbeeld bij natrium) een hydratatieschil vormt. Eiwitvouwing, baseparing in DNA, enzymkinetiek en vele andere biochemische fenomenen in de cel komen tot stand door interacties met watermoleculen.
Biologische betekenis
Water is een van de belangrijkste componenten van cellen. Het cytosol, de intracellulaire grondvloeistof van de cel, bevat de eiwitten en de organische moleculen die nodig zijn om het organisme in leven te houden.
Water is een reactant in vele biochemische reacties. Een noemenswaardig geval zijn de lichtreacties, waarin een watermolecuul wordt gesplitst onder invloed van licht. Het ontstane waterstofion wordt gebruikt voor de opbouw van glucose, het zuurstof komt vrij als bijproduct.
Water biedt structurele ondersteuning via turgor, de druk die het celmembraan tegen de celwand duwt. Bij landplanten zorgt turgordruk voor stevigheid van het plantenlichaam. Water maakt daarnaast celstrekking mogelijk.
Water is het medium voor de overdracht van plantaardige gameten. Bij mossen en veel varens zwemmen de mannelijke gameten, de zaadcellen naar de vrouwelijke gameten, de eicellen door het water; sommige waterplanten geven stuifmeel af onder water.
Water bemiddelt de verspreiding van zaden en vruchten (propagulen), zoals bij de kokosnoot, els of gele lis.
Beweging bij planten kunnen het gevolg zijn van water dat in en uit de bewegende delen diffundeert. Een voorbeeld is de dagelijkse bewegingen van huidmondjes: deze openen en sluiten in reactie op veranderingen in licht en luchtvochtigheid.
Een belangrijk aspect in de evolutie van planten is de bemachtiging van voldoende water Plantenmorfologische kenmerken van planten hangen vaak samen met de beschikbaarheid van water, zoals organismen die groeien in droge, matig droge en natte habitats.
Diffusie en osmose

Vrijwel alle transportprocessen op cellulair en intercellulair niveau berusten op diffusie: de in wezen ongerichte, willekeurige warmtebeweging van deeltjes. Bij verschillen in concentratie leidt diffusie tot een netto verplaatsing van deeltjes van plaatsen met een hoge concentratie naar plaatsen met een lage concentratie. De netto beweging stopt wanneer een dynamisch evenwicht is bereikt.
Osmose is een proces op basis van diffusie waarbij een vloeistof, waarin stoffen zijn opgelost, door een semipermeabel membraan beweegt, dat wel de vloeistof doorlaat maar niet de opgeloste stoffen. In plantenfysiologie is de vloeistof vrijwel altijd water. Het membraan van plantencellen is permeabel voor kleine, lipofiele, hydrofobe en ongeladen moleculen: water, ammoniak, zuurstof, enkele hormonen en neutrale aminozuren. Ze zijn niet permeabel voor grote, hydrofiele en geladen deeltjes zoals eiwitten, suikers en ionen. Voor zulke moleculen bevatten celmembranen gespecialiseerde transporteiwitten. Er zijn naast diffusie en osmose nog twee andere transportprocessen van belang:
- Massabeweging of bulkstroom. De moleculen bewegen van hoge naar lage druk volgens een drukgradiënt. Vaak gaat het om de beweging van water vanaf de wortels naar de bladweefsels.
- Dialyse is een ander gespecialiseerd geval van diffusie. Het is de diffusie van opgeloste stof door een semipermeabel membraan. Als water (het oplosmiddel) uit de cel beweegt in de omgeving, beweegt het osmotisch. Indien het suiker (opgeloste stof) beweegt in de omgeving, is een voorbeeld van dialyse.
De wet van Fick is een veelgebruikte vergelijking die de diffusiesnelheid in verband brengt met de concentratiegradiënt C1 − C2 en de weerstand r. De weerstand is afhankelijk van factoren als afstand, gemiddelde viscositeit en membraanpermeabiliteit. De diffusiesnelheid (of fluxdichtheid, Js in eenheden van mol m2 s1) kan worden uitgedrukt in de vereenvoudigde versie van de wet van Fick:
- .

Deze vergelijking kan gebruikt worden om osmose (diffusie van water) te beschrijven, maar ook voor andere deeltjes die vrij kunnen diffunderen. Verschillende factoren beïnvloeden de diffusiesnelheid:
- Moleculaire bewegingssnelheid: volgens de kinetische theorie zijn deeltjes zoals atomen en moleculen in altijd in beweging bij een temperatuur boven het absolute nulpunt (0 K = −273 °C). Moleculaire beweging is direct evenredig met de temperatuur, en heeft een indirect verband met molecuulgewicht: zwaardere deeltjes bewegen langzamer dan lichtere, kleinere.
- Temperatuurverhoging verhoogt de snelheid van de moleculaire beweging, dus verhoogt van de diffusiesnelheid
- Druk verhoogt de snelheid van moleculen, dus verhoging van de diffusiesnelheid
- Het effect opgeloste stof op de chemische potentiaal van het oplosmiddel. Opgeloste deeltjes verlagen de vrije energie van een oplosmiddel. De kritische factor is het aantal deeltjes, niet de lading of de deeltjesgrootte. De moleculen van het oplosmiddel, zoals water in een biologisch systeem, bewegen van een gebied van hogere molfractie naar een gebied met een lagere molfractie.
Waterpotentiaal
Het waterpotentiaal (Ψw) is een maat voor de energietoestand van water. Dit is een bijzonder belangrijk concept in de plantenfysiologie, omdat het de richting en beweging van het water bepaalt. Bij een verschil in μw aan weerszijden van een membraan zal osmose optreden in de richting van de oplossing met de laagste waterpotentiaal. De vergelijking voor waterpotentiaal moet rekening houden met de factoren die de diffusie van water beïnvloeden:
| = | + | + | ||||
| waterpotentiaal | = | drukpotentiaal | + | osmotisch potentiaal | + | zwaartekrachtspotentiaal |




De waterpotentiaal Ψw is de chemische potentiaal van water (in J mol−1 = MPa) in vergelijking met zuiver water bij dezelfde temperatuur en druk. De grootheid is druk omdat plantencellen onder druk staan (denk aan de celwand) en het gemakkelijker is om te meten. De waterpotentiaal van zuiver water is nul. De waterpotentiaal in intact plantenweefsel is meestal negatief vanwege de grote hoeveelheden opgeloste stoffen in de cellen.[c]
- De drukpotentiaal (Ψp) ontstaat ten gevolge van de drukopbouw in cellen door de celwand. De drukpotentiaal Ψp is meestal positief, maar kan negatief (onderdruk) zijn in het xyleem. Druk kan worden gemeten met een osmometer. Druk wordt gemeten in megapascal (MPa). 1 MPa komt overeen met 10 bar, wat ongeveer 10 atmosfeer is.
- De osmotische potentiaal (Ψs) is de bijdrage als gevolg van opgeloste stoffen. Opgeloste stoffen geven altijd een vermindering van de vrije energie van het water, waardoor hun bijdrage altijd negatief is. De osmotische potentiaal kan gemeten worden door middel van het vriespuntverlaging (opgeloste stoffen verlagen het vriespunt van een vloeistof), grensplasmolyse vast te stellen. De osmotische potentiaal Ψs van een oplossing kan worden berekend met behulp van de Van 't Hoff-vergelijking:
- .

Hierin is i de ionisatieconstante (de van 't Hoff-factor, vaak gelijk aan 1), M de concentratie van de oplossing (of het concentratieverschil), R de molaire gasconstante (8,3144 J mol−1 T−1) en T de temperatuur (in K).
- De matrixpotentiaal (Ψm) is de bijdrage aan de waterpotentiaal als gevolg van de aantrekkingskracht van water voor colloïdale, geladen oppervlakken. Deze is negatief omdat het van water de mogelijkheid om te bewegen beperkt. In grote volumes is de matrixpotentiaal verwaarloosbaar. Het kan echter belangrijk zijn in de bodem, vooral met betrekking tot het wortel-bodem raakvlak. Zo bestaat klei uit kleine, colloïdale gronddeeltjes (kleiner dan 2 µm).
- De zwaartekrachtspotentiaal (Ψg) of gravitatiepotentiaal is de bijdrage van zwaartekracht. Deze is gewoonlijk verwaarloosbaar, tenzij men rekening houdt met de invloed van de zwaartekracht bij transport in hoge bomen.
De verplaatsing van het water door een membraan is een combinatie van diffusie en "bulk flow". Individuele watermoleculen diffunderen door het membraan. Daarnaast heeft het membraan ingebouwde eiwitten die een kanaal of porie vormen waardoor het water kan bewegen. Deze poriën zijn belangrijk voor het transport van watermoleculen door "bulk flow". De zogenaamde aquaporine-eiwitten van de poriën vormen deze transportkanalen voor het water. Water beweegt zich passief en volgt het gradiënt van vrije energie.
Watertransport
Het transport van water is voor de plant van groot belang.[7] In het continuüm van bodem-plant-lucht volgt het water de route van de bodem, via de wortels en de stengel, langs de bladeren tot weer in de lucht. De drijvende kracht voor beweging van het water is de gradiënt in de waterpotentiaal die bestaat tussen de bodem en de lucht. Met andere woorden:
| Route: | bodem | → | wortel | → | stam | → | twijg | → | blad | → | lucht |
| Proces: | {wateropname}
|
→ | {watertransport}
|
→ | {transpiratie}
| ||||||
| Waterpotentiaal: | bodem | > | wortel | > | stam | > | twijg | > | blad | > | lucht |
karakteristieke
waarden (boom) |
gravitatiewater: 0 MPa |
· |
-0,7 MPa |
-2,3 MPa |
-2,5 MPa |
||||||

Overgang van bodem naar plant
De eerste stap bij het watertransport is de overgang van de bodem naar de plant. De bodem is de laag van de aardkorst die door planten beworteld is en waarin zich bodemvormende processen afspelen. De bodem bestaat uit een mengsel van organische stoffen (humus, planten- en dierenresten, mest, reducenten), anorganische materialen (verweerde rots), gassen (zuurstof, kooldioxide, ethyleen) en vloeistof (water). De grondsoort wordt bepaald door de samenstelling, textuur of deeltjesgrootte, en de structuur (met name de bodemverdichting).
Gravitatiewater is water dat afstroomt en niet sterk gebonden is; Ψ = 0 MPa. Veldcapaciteit heeft de grond als die met al het tegen de zwaartekracht in vastgehouden water verzadigd is. Het capillaire water is het water dat wordt vastgehouden door capillaire werking of het water bij veldcapaciteit; Ψ = −0,015 MPa. Als het vochtgehalte van de bodem lager is dan waarop planten kunnen groeien treedt permanente verwelking op, gewoonlijk bij Ψ = −1,5 MPa. De bodemwaterpotentiaal is een functie van osmotische potentiaal (in zoute gronden wijkt deze sterker af van 0) en vooral van de druk of matrixpotentiaal (adhesie, onderdruk die veroorzaakt wordt door de aantrekkingskracht van colloïdale deeltjes). De druk in de bodem kan worden berekend uit de vergelijking:
- .

Hierin is de oppervlaktespanning (7,28 · 10−8 MPa) en de boogstraal van de meniscus. Verplaatsing van het water in de bodem is vooral te danken aan bulkstroom (massabeweging) als gevolg van drukgradiënten en aan enige diffusie.


Routes

Het meeste water wordt aan het uiteinde van de wortel opgenomen, omdat daar de nog levende wortelharen zitten. Hoe verder van het punt, hoe minder water wordt opgenomen door de wortel. Dat komt ruwweg overeen met het gebied van de wortel dat verkurkt.
Er zijn drie routes die het water kan volgen:
- apoplastisch: het water volgt een apoplastisch route (door de "niet levende" delen van de plant) van de bodem door de schors. Wel moet het in de centrale cilinder komen via de symplast (de "levende" delen van de plant) vanwege de ondoorlaatbare bandjes van Caspary. Eenmaal binnen lekt het uit naar de apoplast (xyleem), waar het wordt getransporteerd naar de top van de plant. Dit blijkt de belangrijkste manier van transport.
- symplastisch door de membranen (transmembraan); met andere woorden, het water beweegt van cel tot cel dwars door de membranen,
- symplastisch door de plasmodesmata: het water beweegt van cel tot cel door de stippels.
Xyleemtransport
Xyleem is het weefsel dat dient voor watertransport, wat bewezen kan worden door onderzoek met tracers. Hierbij wordt xyleem voorzien met kleurstoffen. Een voorbeeld is de klassieke experiment met een stengel van bleekselderij die in een kleurstof wordt geplaatst. Het transport van de kleurstof door de vaatbundels is dan duidelijk zichtbaar. Het transport vindt plaats in de tracheeën en tracheïden. Er zijn vier belangrijke types van cellen:
- Tracheïden: lange, taps toelopende uiteinden, dikke secundaire wand;
- Trachee-elementen: een trachee bestaat uit meerdere cellen die onderling verbonden zijn door openingen over de volledige diameter van de cel;
- Sclereïden (vezelcellen): lang en slanke cellen met een dikke secundaire wand, meestal voor ondersteuning;
- Parenchymcellen: levende, dunwandige cellen die zetmeel opslaan en algemene fysiologische functies vervullen
-
 Microscopische doorsnede van het vaatweefsel van een zonnebloem (Helianthus). De grote witte cirkels zijn individuele xyleemvaten of tracheïden.
Microscopische doorsnede van het vaatweefsel van een zonnebloem (Helianthus). De grote witte cirkels zijn individuele xyleemvaten of tracheïden. -
 Microscopische doorsnede van de stengel van een wonderboom (Ricinus). Centraal in de stengel ligt het protoxyleem met naar buiten het jongere metaxyleem.
Microscopische doorsnede van de stengel van een wonderboom (Ricinus). Centraal in de stengel ligt het protoxyleem met naar buiten het jongere metaxyleem.


De primaire cellen voor watertransport zijn tracheïden en tracheeën. Naaktzadigen hebben uitsluitend tracheïden terwijl bedektzadigen over beide typen beschikken en in de eerste plaats afhankelijk zijn van tracheeën voor het watertransport. Zowel tracheïden en tracheeën zijn voorzien van zogenaamde stippels: ronde verdunning in de celwand waardoor de uitwisseling van stoffen tussen cellen gemakkelijker kan plaatsvinden.
Mechanisme van watertransport
Er zijn verschillende mogelijke mechanismen denkbaar voor het watertransport van beneden naar boven in de plant of boom: een pomp, capillaire werking en cohesie in combinatie met onderdruk in de vaatbundels.
- Voor een pomp die zou zorgen voor druk vanaf beneden is er anatomisch geen orgaan te vinden. Daarnaast wordt picrinezuur omhoog getransporteerd, terwijl dat toch de cellen doodt. Ook de worteldruk alleen is slechts 0,2-0,3 MPa, maar een druk van ten minste 3 MPa is nodig om water naar de top van hoge bomen te transporteren.
- Capillaire werking is niet verantwoordelijk voor de verplaatsing van het water omdat de tracheeën te breed zijn om het transport voldoende hoog te ondersteunen.
- Volgens de cohesie-onderdruk theorie wordt het water opgezogen door de plant door de kracht van de verdamping (evaporatie) aan het bladoppervlak. Bij de verdamping van watermoleculen bij de huidmondjes worden andere watermoleculen aangezogen door de cohesieve en adhesieve eigenschappen van water.
Wil cohesie-onderdruk theorie kloppen, dan moet het watertransport in planten aan de volgende criteria voldoen:
- Het systeem moet weinig weerstand vormen. De tracheeën en tracheïden zijn bij volwassenheid hol: ze hebben geen plasma en zijn dood.
- Tussen de bladeren en de bodem moeten de waterkolommen aaneensluitend zijn. De tracheïden en tracheeën vormen een continue waterkolom.
Cavitatie is het breken van de waterkolom door onderdruk (het vacuüm trekken) waardoor lucht uit de oplossing komt. Als er lacunes of luchtbellen zijn ontstaan, moet de waterkolom worden omgelegd rond deze bellen. Als cavitatie optreedt reageert de plant door:- het transporteren van water rond de geblokkeerde cel
- opnieuw oplossen van de luchtbel, wat meestal 's nachts plaatsvindt
- vorming van nieuwe xyleemcellen: xyleem is vervangbaar. Alleen de meest recente gegroeide cellen van de laatste seizoenen zijn functioneel. Het overige hout in een boom is niet functioneel door cavitatie of omdat ze gevuld zijn met afvalstoffen. Verder neemt men aan dat hofstippels de functie hebben het transport van de luchtbellen te stoppen uit een cel waar cavitatie is opgetreden en daardoor het effect van cavitatie te isoleren.
- Er moet voldoende trekkracht te zijn. Ook al is ongeveer 3 MPa nodig om water naar de top van een hoge boom te bewegen, het waterpotentiaalgradiënt van bodem naar lucht aanzienlijk steiler (in de grootteorde van 100 MPa.)
- Het xyleem staat onder een onderdruk. Verscheidene lijnen van bewijsmateriaal ondersteunen deze voorspelling:
- snijd een stengel door en de waterkolom breekt in de top en het water hoopt zich op bij het snijvlak aan de onderkant
- terwijl de boom transpireert wordt de diameter van de boom gemeten met een dendrometer. Gedurende de dag is de diameter van de stam het kleinste en 's nachts het grootste.
- aanprikken van het xyleem van een actief ademende boom met een ijspriem kan resulteren in een sissend geluid als de lucht wordt aangezogen in de steel
- kleurstof-oplossingen worden snel opgezogen in een boomstam als zij met een mes worden aangeprikt, de kleurstof wordt vervolgens getransporteerd in zowel opwaartse en neerwaartse richting. Aangezien de druk in de stengel lager is dan atmosferische wordt de kleurstofoplossing snel aangezogen.
- De treksterkte van de waterkolom moet de zuigkracht kunnen weerstaan. Met andere woorden, de waterkolom mag niet breken als eraan wordt getrokken. Als het water in de waterkolom omhoog wordt getrokken ontstaat er een weerstand. Water heeft een zeer hoge treksterkte. Wanneer de onderdruk te groot is, veroorzaakt dit dat de gassen uit de oplossing komen en een dampbel vormen (cavitatie). Naarmate de houtvaten wijder zijn, neemt de kans op cavitatie toe. Cavitatie kan worden gehoord met behulp van gevoelige microfoons op de plant, waarbij kleine plopgeluiden zijn te horen.
- Tracheïden en tracheeën moeten in staat zijn om onderdruk te weerstaan zonder imploderen. Dit is de reden dat ze dikke celwanden hebben met spiraalvormige verdikkingen. Hierdoor is hout zeer hard.
Floëemtransport

Voornamelijk in het floëem vindt het transport van stoffen, translocatie plaats, maar het kan ook in het xyleem. Water en opgeloste ionen zijn de belangrijkste stoffen in tracheeën en tracheïden. Deze materialen worden vervoerd via transpiratie stroom. Xyleemsap kan ook organische materialen vervoeren, gewoonlijk in relatief lage concentraties (met als opvallende uitzondering esdoornsap dat in het voorjaar bestaat uit 2% of meer sacharose). Stoffen bewegen met verschillende snelheden, onder andere afhankelijk van matrixeffecten, metabole behoeften.[8]
Floëem is om verschillende redenen moeilijk te bestuderen omdat de transportcellen en -weefsels microscopisch klein zijn (in vergelijking met de transportstructuren in dieren); er is een zeer snelle respons van het floëem om verwonding omdat de inhoud onder druk staat; het transport bij planten vindt intracellulair plaats (daarentegen extracellulair bij dieren) en de transportcellen zijn levend.
Floëem is het primaire transportweefsel voor producten van de fotosynthese (fotoassimilaten, organische stoffen). Bladluizen prikken met hun steeksnuit de direct de floëemcellen aan. De bladluizen zuigen niet, maar omdat de floëemcellen onder druk staan, krijgen de bladluizen soms zo veel vloeistof binnen, dat het er aan de achterkant weer uit komt als honingdauw.
Samenstelling van het floëemsap:
- 16−25% koolhydraten (zoals sacharose, stachyose, raffinose),
- 0,04−4% amines en amiden (zoals asparagine, glutamine, asparaginezuur, ureum, citrulline en allantoïne),
- ATP, plantenhormonen, suikeralcoholen zoals sorbitol en mannitol, en een verscheidenheid van andere organische stoffen,
- Anorganische stoffen, waaronder magnesium en kalium.
Floëem transporteert organische materialen van de plaats van de productie ("source") naar een plaats waar er behoefte aan is ("sink"). De richting van transport is gewoonlijk naar beneden: van de primaire bron, de bladeren naar de belangrijkste "sink", de wortels. De snelheid van het transport in het floëem bedraagt gemiddeld ongeveer 30 cm per uur of nog sneller.
Gaswisseling en transpiratie
Transpiratie is de verdamping van water uit oppervlakte van een plant. Lucht heeft een zeer hoog vermogen om het water vast te houden. Er is een zeer steile gradiënt in de waterpotentiaal van bodem naar de lucht. De plant plaatst zichzelf tussen die twee en maakt zo gebruik van passief transport. Evapotranspiratie is de verdamping van water uit een plant en de grond (met inbegrip abiotische omgeving). Planten verliezen veel water door transpiratie.

Diffusie van water en kooldioxide
Planten hebben een groot oppervlak nodig voor een efficiënte gasuitwisseling. Daartoe bestaan bladeren voor een groot deel uit een sponsachtig weefsel met intracellulaire holten: het chlorenchym. De bovenkant van het blad is bedekt met een voor water ondoordringbare cutinelaag: de cuticula. Verder is het blad rijkelijk voorzien van huidmondjes (stomata). Deze strategieën minimaliseren risico’s op uitdroging.
De plaats van de absorberende oppervlakken inwendig in het organisme (vooral in de bladeren) om verdroging te verminderen vormt een probleem bij het verkrijgen van de gassen aan de absorptie-oppervlak. Hoewel planten geen pompmechanisme voor het verplaatsen van gassen hebben (zoals dieren), kan door de zeer geringe dikte van de bladeren de diffusie (en in mindere mate bulk flow) een grote rol spelen.
In de lucht heeft CO2 een lage concentratie, in het blad nauwelijks minder. In het blad is de lucht bijna verzadigd met waterdamp, in de lucht meestal bij lange na niet. Bij een relatieve luchtvochtigheid van 50% is de waterpotentiaal-gradiënt 100 MPa (of 32 mmol mol−1). Gebaseerd op gradiënt alleen al heeft water ongeveer 100× meer neiging het blad uit te diffunderen dan kooldioxide het blad in te diffunderen. Diffusiesnelheid is omgekeerd evenredig aan het molecuulgewicht. Dus hoe zwaarder het molecuul des te langzamer het zal diffunderen.
Huidmondjes

Huidmondjes zijn van groot belang bij de regulering van de verdamping en de opname van kooldioxide.[9] Een huidmondje bestaat uit sluitcellen, hulpcellen, de substomatale holte, een cuticula, een richel of lip op de sluitcellen. De epidermale hulpcellen zijn gespecialiseerde cellen die afwijkend kunnen zijn van de andere epidermiscellen. De functie van de richel is het voorkomen dat vloeibaar water in de opening sijpelt.
Een laag met cutine bedekt het grootste deel van de cellen in de substomatal holte, alleen het gebied bij de werkelijke opening is vrij van de cuticula. Het meeste waterverlies vindt plaats in dit gebied. Er is een waterdichte bekleding op het absorberende bladoppervlak dat rijkelijk voorzien is van een eindeloze hoeveelheid openingen. Er zijn tot wel 1000 stomata per mm2, die gezamenlijk maar liefst 2 tot 3% van het totale bladoppervlak bezetten. Een huidmondjes is slechts 14 μm in diameter.
De huidmondjes vormen ideale structuren voor de regulering van de gasuitwisseling. Kleinere huidmondjes zijn efficiënter dan grotere door het randeffect. De mate van openen en sluiten van de poriën wordt door de plant gereguleerd als antwoord op het milieu. Elke factor die de fotosynthese of algemene waterkwaliteit van de plant beïnvloedt, beïnvloedt de werking van de sluitcellen.
Sluitcellen openen als door het osmotische binnendringen van water. Op zijn beurt verhoogt dit de turgor (waterdruk) in de sluitcellen waardoor ze zich strekken. De radiale oriëntatie van cellulose microfibrillen voorkomt toename van de omtrek. De sluitcellen zijn aan hun einde aan elkaar bevestigd en hun binnenwand is dikker; hierdoor krommen zich de sluitcellen en opent zich het huidmondje.
Groei, ontwikkeling & hormonen
Plantenhormonen
Planten bezitten een goed ontwikkeld systeem van de chemische boodschappers die groei en reacties in de ontwikkeling induceren (remmen of bevorderen). Deze chemische boodschappers worden aangeduid als "hormonen".[10][11]
Plantenhormonen zijn kleine organische verbindingen, die gesynthetiseerd worden door de plant. Ze zijn werkzaam in lage concentratie (<10−6), ze bevorderen of belemmeren de groei en ontwikkelingsreacties en vertonen vaak een scheiding van de plaats van productie en de plaats van de werking. Er zijn relatief weinig plantenhormonen, elk roept een verscheidenheid van reacties op en ze werken vaak samen met andere hormonen.
Er zijn vijf belangrijke groepen, gebaseerd op chemische structuur. Met uitzondering van de laatste twee omvat elke groep een aantal verwante verbindingen. Deze groepen zijn auxines, gibberellines, cytokininen, abscisinezuur en etheen (ethyleen).
Daarnaast zijn er tal van andere "plantenhormonen" zoals brassinosteroïden, oligosacharides, polyamines, jasmonaten, salicylzuur en systemin.
Door inwerking op het doelweefsels activeren hormonen een receptor:
| hormoon → doelweefsel/cel → receptor → signaalversterking → reactie |
Dus om een reactie te doen optreden moet het hormoon in voldoende mate aanwezig zijn; moet het doelweefsel gevoelig zijn of gemaakt (gesensibiliseerd) voor het hormoon; moet het doelweefsel het hormoon herkennen en moet er een receptor zijn waaraan het hormoon zich kan binden; moet de binding van het hormoon en receptor een verandering in de receptor initiëren (amplificatie) en dan initieert de geactiveerde receptor een fysiologische respons.
Auxines
Auxine is een algemene naam voor een groep van hormonen die betrokken zijn bij groeiresponsen. De term auxine is afgeleid van het Griekse woord "toenemen of groeien". Auxine was de eerst ontdekte groep van plantenhormonen.[12]
De belangrijkste auxine in planten indool-3-azijnzuur (IAA). Andere auxines die zijn geïsoleerd uit planten worden waarschijnlijk in vivo omgezet IAA.
Auxinen treden net als andere hormonen op in een vrije of geconjugeerde (gebonden aan suikers, alcoholen of andere moleculen) vorm. Tot 98% van de auxine gebonden met inositol, acetyl-CoA of glycosiden.
IAA wordt niet getransporteerd door de transpiratiestroom of door het floëemtransport, want de transportsnelheid is te langzaam. De transportsnelheid komt overeen met die van diffusie.
Gibberellines
Gibberellines zijn een groep van plantenhormonen.[13] De naam gibberelline is afgeleid van de door Eiichi Kurosawa ontdekte schimmel Gibberella fujikuroi. De door de schimmel gevormde gibberelline veroorzaakte een sterke plantengroei waardoor slappe planten met lange internodiën ontstonden.
Cytokininen
Cytokinines[14] zijn een groep celdelingshormonen, die ook van invloed op de celgroei en celdifferentiatie. Ze worden in de worteltoppen gevormd en bevorderen de groei van de zijscheuten en dus de doorbreking van de apicale dominantie. Verder gaan deze hormonen de veroudering tegen.
Abscisinezuur
Abscisinezuur[15] is een plantenhormoon dat zorgt voor tolerantie tegen stress door vocht- of voedseltekort door de reactie van de plant op stress naar de specifieke organen over te brengen. Het is een sleutelsignaal dat de mate van opening van de huidmondjes regelt en is belangrijk bij het in rust brengen van zaden en knoppen. Ook is het samen met andere signaalstoffen betrokken bij de reactie van de plant op verwonding en aantasting door ziekteverwekkers. Elke plantencel kan abscisinezuur synthetiseren.
Etheen
Etheen (of ethyleen)[16] heeft verschillende effecten. Een ervan is dat etheen wordt geproduceerd door sommige rijpende vruchten, de zogenaamde climacterische vruchten. Voorbeelden van climacterische vruchten zijn appels, passievruchten, bananen, kaki's, avocados en tomaten. Etheen kan ook gebruikt worden voor de kiemremming bij uien.
Voor bloeminductie bij bromelia's wordt meestal gebruikgemaakt van ethyn[17], maar ook ethyleen kan hiervoor gebruikt worden.
Minerale voeding
De minerale samenstelling van planten is gekenmerkt door:[18]
- 90% of meer van de plant bestaat uit de elementen koolstof (C), waterstof (H), zuurstof (O) en stikstof (N);
- in kleinere hoeveelheden worden er in planten meer dan 60 andere elementen gevonden, zoals goud (Au), arseen (As), kwik (Hg), lood (Pb) en uranium (U);
- bepaalde weefsels hopen sommige elementen op, bijvoorbeeld jonge weefsels hebben hogere gehalten aan stikstof (N), fosfor (P) en kalium (K);
- verschillende plantensoorten hebben een vergelijkbare chemische samenstelling;
- niet alle in een plant voorkomende elementen zijn essentieel, en sommige elementen kunnen niet selectief geabsorbeerd worden
Sommige elementen worden opgenomen uit de lucht, met name C en, gedeeltelijk, O (via CO2 en O2), de andere benodigde elementen worden door de wortels uit de grond geabsorbeerd.
Functies, waarbij de elementen betrokken zijn:
- C, H, O, N en S zijn nodig omdat ze de bouwstenen zijn van de moleculen van het leven;
- P en B (en Si) zijn betrokken bij energieoverdracht-reacties;
- K, Na, Mg, Ca, Mn en Cl hebben verschillende functies, waaronder de handhaving osmotische concentratie en structuren van enzymen;
- Fe, Cu, Zn en Mo zijn nodig in prostetische groepen voor de reacties met elektronenoverdracht.
Stikstof, fosfor en kalium zijn meestal de gebruikelijke beperkende elementen, omdat ze óf nodig zijn in de hoogste concentratie, óf het minst vaak in voldoende hoeveelheden worden geleverd door de bodem (of een ander groeimedium). Bij de plantenteelt is er hierom een behoefte aan meststoffen voor de gewassen.
| De belangrijkste elementen in de bodem en in de plant | ||||
|---|---|---|---|---|
| Belangrijke elementen |
Voorkomen in bodem |
Macro-element, sporenelement |
Potentieel giftig |
Opmerkingen |
| Waterstof (H als H2O) | + | + | cellulose, zetmeel en andere organische verbindingen | |
| Boor (B als HBO3) | sporenelement | † | ||
| Koolstof (C als CO2) | + | + | cellulose, zetmeel en andere organische verbindingen | |
| Stikstof (N als NO3–, NH4+) | + | nucleïnezuren, proteïnen, hormonen, etc. | ||
| Zuurstof (O als O2, H2O) | + | + | cellulose, zetmeel en andere organische verbindingen | |
| Fluor (F als F−) | † | |||
| Natrium (Na als Na+) | ||||
| Magnesium (Mg als Mg2+) | + | + | essentiële bestanddeel van chlorofyl | |
| Aluminium (Al) | + | † | ||
| Silicium (Si) | + | |||
| Fosfor (P als H2PO4−) | + | nucleïnezuren, fosfolipiden, ATP | ||
| Zwavel (S als SO42−) | + | bestanddeel van proteïnen | ||
| Chloor (Cl als Cl−) | sporenelement | |||
| Kalium (K als K+) | + | + | cofactor in eiwitsynthese, waterhuishouding, etc. | |
| Calcium (Ca als Ca2+) | + | + | Synthese van membranen en stabilisatie | |
| Vanadium (V) | † | |||
| Chroom (Cr) | † | |||
| Mangaan (Mn als Mn2+) | sporenelement | † | ||
| IJzer (Fe als Fe2+, Fe3+) | + | sporenelement | ||
| Kobalt (Co) | † | |||
| Nikkel (Ni als Ni2+) | sporenelement | † | ||
| Koper (Cu als Cu+) | sporenelement | † | ||
| Zink (Zn als Zn2+) | sporenelement | † | ||
| Arseen (As) | † | |||
| Seleen (Se) | † | |||
| Broom (Br) | † | |||
| Strontium (Sr) | † | |||
| Molybdeen (Mo als MoO42−) | sporenelement | † | ||
| Cadmium (Cd) | † | |||
| Tin (Sn) | † | |||
| Barium (Ba) | † | |||
| Kwik (Hg) | † | |||
| Lood (Pb) | † | |||
Plantenrespons op het milieu
- Aanpassing van planten aan hun groeiplaats.[19] Planten kunnen zich niet verplaatsen, maar kunnen zich wel min of meer aanpassen aan hun groeiplaats door:
- het verspreide zaad al of niet te laten kiemen (kiemrust, lichtkiemer)
- gravitropie, groei van planten beïnvloed door de zwaartekracht.
- maximale aanpassing aan hun milieu door o.a. fototropie, en door zich te handhaven op voor andere planten giftige gronden (zinkviooltje).
- genetische aanpassing (survival of the fittest=overleven van de best aan de omgeving aangepaste).
- Fytochroom is een fotoreceptor-eiwit dat betrokken is bij het waarnemen van licht door de plant: het eiwit werkt als een licht-sensor. Fytochroom B1 absorbeert het rode licht van 660 nm, terwijl fytochroom B2 het verrood licht van 730 nm absorbeert. Daglicht bestaat voor een groot gedeelte uit rood licht, en er is dan vooral het fytochroom B2 aanwezig. Gedurende de nacht verandert het fytochroom B2 in fytochroom B1. Fytochroom B2 gaat de strekking van zaailingen tegen, omdat de kieming in de grond plaatsvindt en dus in het donker gebeurt. Komt de zaailing boven de grond dan moet de strekkingsgroei stoppen. Ook bladgroei, chlorofyl aanmaak en het rechtop laten groeien van het epicotyl of hypocotyl van zaailingen worden beïnvloed door het fytochroom.
- Bloeiperiode, zoals bloei bijvoorbeeld in het voorjaar (lange-dagplanten of bij het bereiken van een bepaalde koude- of warmtesom), zomer of winter.
- Het wortelstelsel wordt door de plant aangepast aan de groeiomstandigheden. Onder droge omstandigheden ontstaat er een uitgebreid wortelstelsel, waarbij de wortels met het dalen van de grondwaterstand ook dieper de grond ingaan. Plantenwortels kunnen ook stoffen afgeven die remmend werken op de groei van andere plantensoorten. Als een deel van de bovengrondse delen afgevreten of beschadigd raken 'probeert' de plant dit zo snel als mogelijk te herstellen, door het verminderen of stoppen van de wortelgroei en soms zelfs gedeeltelijk afsterven van het wortelstelsel, zodat alle energie in hernieuwde stengel en bladvorming kan gaan zitten. Bij beschadiging of gedeeltelijk afsterven van het wortelstelsel treedt het omgekeerde op, en sterft het bovengrondse deel van de plant gedeeltelijk af.
Prikkelbaarheid
Planten zijn, net als andere organismen, in staat te reageren op veranderingen in hun omgeving, op verscheidene manieren.
- Planten kunnen reageren door diverse bewegingen (tropie, nastie en taxie) op meestal onvoorspelbare, kortdurende schommelingen in hun omgeving, zoals plotselinge veranderingen in neerslag, temperatuur of licht. Zo zijn er planten waarvan de bloemen sluiten bij een regenbui.
- De vorm van planten reageert op hun omgeving. Licht is een van de belangrijkste omgevingsfactoren voor de ontwikkeling van planten. Dit verschijnsel heet fotomorfogenese; een klassiek voorbeeld is etiolering: bij lichtgebrek groeien planten dun en lang uit en blijven gelig groen.
Verdediging
Zowel de fysische omgeving als het biotische milieu dragen risico's in zich voor het welzijn van de planten. Om zich in stand te houden moet de plant zich tegen deze omgevingsfactoren verdedigen. Tot de fysische gevaren behoren wind, water (overstroming), droogte en kou (winter). Hiertegen beschermen planten zich door kiemrust, veroudering, en - tegen de meer voorspelbare, seizoensgebonden gevaren - zelfs door dood te gaan (eenjarige planten). De bladverliezende levenswijze is een vorm van adaptatie: een periodiek terugkerende reactie op seizoensgebonden, ongunstige omstandigheden.
Tot de biologische gevaren behoren de herbivoren of planteneters, schimmels, insecten en concurrenten (de andere planten). Ter verdediging hebben planten verschillende aanpassingen, met name in morfologie (doornen en stekels, haren en een dikke cuticula op de epidermis), chemie (giftige of onverteerbare secundaire plantenstoffen), mimicry en signaalstoffen. Ook kunnen planten communiceren.
Metabolisme
Stofwisseling of metabolisme is het geheel van biochemische processen in organismen. Er kan onderscheid gemaakt worden tussen anabolisme (de opbouw van stoffen met gebruik van energie), en katabolisme (de afbraak van complexere stoffen waarbij energie weer vrijkomt.) Het evenwicht tussen anabolisme en katabolisme wordt onder andere bepaald door het circadiaan ritme. Enzymen spelen bij de omzettingen in de cel een centrale rol.
Anabolisme
Anabolisme is de benaming voor een reeks metabole routes waardoor uit kleinere eenheden moleculen worden gevormd.[20] Een dergelijk metabolisch proces kan zich afspelen op het niveau van individuele cellen, organen of hele organismen. De meeste anabolische processen worden aangedreven door adenosinetrifosfaat.
Katabolisme
Katabolisme is de benaming voor verschillende manieren van stofwisseling in een cel, waarbij grote moleculen, zoals polysacharide, lipiden, nucleïnezuren en eiwitten worden afgebroken in kleinere moleculen (respectievelijk monosachariden, vetzuren, nucleotiden en aminozuren). Bij deze reactie komt energie vrij, waardoor de reactie een vorm is van dissimilatie. De monomeren die als gevolg van het uiteenvallen van polymeren ontstaan worden door cellen gebruikt om nieuwe polymeren te vormen, dan wel om de gevormde monomeren nog verder af te breken, waarbij onder andere koolstofdioxide wordt gevormd. Een voorbeeld van een katabool proces is de citroenzuurcyclus.
Fotosynthese
Fotosynthese is het proces waarin lichtenergie wordt gebruikt om koolstofdioxide om te zetten in koolhydraten. Het proces komt voor in planten en sommige bacteriën. De meeste fotosynthetiserende organismen gebruiken naast koolstofdioxide ook water om deze suikers te maken.
Fotosynthese in planten: C3, C4 en CAM

Bij planten zijn op hoger niveau twee hoofdtypen van fotosynthese te onderscheiden, de zogenaamde C3- en C4-planten. Alle planten hebben het C3-systeem, kooldioxide wordt via tussenproducten met drie koolstofatomen vastgelegd (koolstofassimilatie) en uiteindelijk in glucose omgezet, waarbij de energie wordt geleverd door licht.
Dat fotonen in staat zijn om chemische energie te veroorzaken wordt mogelijk gemaakt door het fotosysteem I en fotosysteem II dat aanwezig is in de chloroplasten die in de thylakoïden pigmenten bevatten zoals chlorofyl. Dit systeem vangt de fotonen en brengt deze van een hoger naar een lager energieniveau. Daarbij komt energie vrij die wordt gebruikt in de Calvincyclus voor de vorming van glucose, sacharose, zetmeel en cellulose.
Er zijn echter planten uit tropische gebieden, zoals mais, die als eerste stap CO2 vastleggen in een verbinding met 4 koolstofatomen, de reden waarom ze C4-planten worden genoemd. De C4-planten doen dit volgens verschillende systemen. De affiniteit van het bindende enzym bij C4-planten voor CO2 is namelijk veel groter. Na deze initiële C4-bindingsstap wordt het CO2 in de plant weer van oxaalacetaat afgesplitst waarbij CO2 en malaat ontstaan en gaat het reactiepad via de normale Calvincyclus verder.
Een andere vorm van fotosynthese is CAM. Dit is een afkorting van 'Crassulacean Acid Metabolism'. Hierbij is de vervolgketen niet ruimtelijk van de initiële CO2-fixatie gescheiden, maar in de tijd. Om namelijk kooldioxide te vangen moeten de huidmondjes van een plant open staan waardoor waterverlies optreedt door verdamping. Bij woestijnplanten met CAM-fotosynthese, veelal succulenten, kan kooldioxide 's nachts worden vastgelegd in malaat of isocitraat en deze verbindingen kunnen dan overdag met behulp van lichtenergie verder worden verwerkt zonder dat de huidmondjes open staan.
De chemische bruto reactie voor fotosynthese is:
12 H2O + 6 CO2 → C6H12O6 + 6 H2O + 6 O2 12 water + 6 kooldioxide → glucose + 6 water + 6 dizuurstof
Fotosynthese begint wanneer in fotosysteem II een chlorofyl pigment een foton (lichtdeeltje) absorbeert. In de Calvincyclus (donkerreactie), die plaatsvindt in de chloroplasten, wordt de door de lichtreactie vastgelegde energie in adenosinetrifosfaat (ATP) gebruikt voor de vorming van glucose uit koolstofdioxide.
De netto reactie van de lichtreactie is:
De netto reactie van de donkerreactie is:
6 CO2 + 12 NADPH2 + 12 ATP → C6H12O6 + 12 NADP + 6 H2O + 12 ADP + 12 Pi 6 kooldioxide + 12 NADPH2 + 12 adenosinetrifosfaat → glucose + 12 NADP + 6 water + 12 ADP + 12 fosfaat